El nudo primitivo (o nodo primitivo ) es el organizador de la gastrulación en el embrión de vertebrado.
Diversidad [ editar ]
- En las aves , se conoce como " nodo de Hensen ", y lleva el nombre de su descubridor Victor Hensen .
- En los anfibios , se conoce como " organizador de Spemann ", y lleva el nombre de Hans Spemann (quien, con Mangold , identificó por primera vez al organizador en 1924. [1] )
Desarrollo [ editar ]
En el desarrollo de los pollitos, el nudo primitivo comienza como un nudo regional de células que se forma en el blastodisc inmediatamente anterior a donde la capa externa de células comenzará a migrar hacia adentro, un área conocida como la línea primitiva , que está involucrada con la hoz de Koller . Posterior al nódulo se encuentra el pozo primitivo, donde las células del epiblasto (la capa superior de las células embrionarias) comienzan a invaginar inicialmente. Esta invaginación se expande posteriormente en el surco primitivo a medida que las capas de células continúan moviéndose hacia el espacio entre las células embrionarias y la yema. Esto diferencia al embrión en las tres capas germinales.- endodermo, mesodermo y ectodermo. El nudo primitivo migra hacia atrás a medida que avanza la gastrulación, y finalmente se absorbe en el brote de la cola.
Modelo predeterminado [ editar ]
Las células del nodo primitivo secretan muchas señales celulares esenciales para la diferenciación neural. Después de la gastrulación, el embrión en desarrollo se divide en ectodermo, mesodermo y endodermo. El ectodermo da lugar al tejido epitelial y neural, siendo el tejido neuronal el destino celular predeterminado. Las proteínas morfogenéticas óseas (BMP) suprimen la diferenciación neural y promueven el crecimiento epitelial. Por lo tanto, el nodo primitivo (el labio dorsal del blastoporo) secreta antagonistas de BMP, incluidos noggin , cordina y follistatina .
El pozo primitivo es una depresión en el centro del nudo primitivo , que se conecta a la notocorda . Consiste en parte de la racha primitiva .
| Hoyo primitivo | |
|---|---|

Vista superficial del embrión de Hylobates concolor . El amnios se ha abierto para exponer el disco embrionario . (El hoyo primitivo no está etiquetado, pero la región es visible).
|
| Rasgo primitivo | |
|---|---|

| |
| Detalles | |
| Etapa Carnegie | 6b |
| Dias | 15 |
| Identificadores | |
| latín | linea primitiva |
| Malla | D054240 |
| Terminologia anatomica | |
La veta primitiva es una estructura que se forma en la blástuladurante las primeras etapas del desarrollo embrionario de aves , reptiles y mamíferos . Se forma en la cara dorsal (posterior) del embrión en desarrollo, hacia el extremo caudal o posterior.
La presencia de la veta primitiva establecerá la simetría bilateral , determinará el sitio de gastrulación e iniciará la formación de la capa germinal . Para formar la línea, los reptiles, las aves y los mamíferos organizan las células mesenquimales a lo largo de la línea media prospectiva, estableciendo el segundo eje embrionario, así como el lugar donde las células ingresarán y migrarán durante el proceso de gastrulación y formación de la capa germinal. [1] La línea primitiva se extiende a través de esta línea media y crea los ejes del cuerpo izquierdo-derecho y craneal-caudal, [2] y marca el comienzo de la gastrulación. [3] Este proceso implica la entrada de mesodermoprogenitores y su migración a su posición final, [2] [4]donde se diferenciarán en la capa germinal del mesodermo [1] que, junto con las capas germinales de endodermo y ectodermo, darán lugar a todos los tejidos del organismo adulto.
Componentes [ editar ]
Dado que el embrión de pollo se puede manipular fácilmente, la mayor parte de nuestro conocimiento sobre la veta primitiva proviene de estudios aviarios. La zona marginal de un embrión de pollo contiene células que contribuirán a la raya. [4] Esta región tiene un gradiente anterior-posterior definido en su capacidad para inducir la línea primitiva, con el extremo posterior con el potencial más alto. [5]
El epiblasto , un blastodisc de capa epitelial única , es la fuente de todo el material embrionario en amniotas [1] y algunas de sus células darán lugar a la línea primitiva. [4] Todas las células en el epiblasto pueden responder a las señales de la zona marginal, [1] pero una vez que una región determinada es inducida por estas señales y experimenta una formación de estrías, las células restantes en el epiblasto ya no responden a estas señales inductivas y prevenir la formación de otra racha. [5]
Subyacente al epiblasto se encuentra el hipoblasto , donde se origina el tejido extraembrionario. [4] En el pollito, la ausencia del hipoblasto produce múltiples rayas, [6] lo que sugiere que su presencia es importante para regular la formación de una sola línea primitiva. En ratones, esta estructura se conoce como endodermo visceral anterior (AVE). [6]
Movimientos celulares [ editar ]
La formación de la veta primitiva en el blastocisto implica el movimiento coordinado y la reorganización de las células en el epiblasto. Incluso antes de que la racha sea visible, las células epiblast han comenzado a moverse. [7] Dos flujos de células contrarrotativos se encuentran en el extremo posterior, donde se forma la línea. [7] Hay poco movimiento en el centro de estos flujos, mientras que el mayor movimiento se observa en la periferia de los vórtices. [3] El movimiento Polonesa es clave para la formación de la racha primitiva. Células superpuestas de la hoz de Kolleren el extremo posterior del embrión de pollo, muévase hacia la línea media, encuentre y cambie de dirección hacia el centro del epiblasto. Las células de la zona marginal posterior lateral reemplazan aquellas células que salieron de la Hoz de Koller al reunirse en el centro de esta región, cambiando de dirección y extendiéndose anteriormente. [4] [8] A medida que estas células se mueven y se concentran en el extremo posterior del embrión, la veta experimenta una transición de la lámina epitelial de una a varias capas que la convierte en una estructura macroscópicamente visible. [4] Varios mecanismos, incluida la división celularorientada , la intercalación célula-célula y el movimiento quimiotáctico de células, [4] Se han propuesto explicar la naturaleza de los movimientos celulares necesarios para formar la línea primitiva.
Formación [ editar ]
La formación de la racha primitiva se basa en una red compleja de vías de señalización que trabajan juntas para garantizar que este proceso esté altamente regulado. Para este proceso se requiere la activación de varios factores secretados (Vg1, Nodal , Wnt8C , FGF8 y Chordin ) y factores de transcripción ( Brachyury y Goosecoid) adyacentes al sitio de formación de la raya. [9] [10] [11] [12] [13] Además, estructuras como el hipoblasto también juegan un papel importante en la regulación de la formación de rayas. La eliminación del hipoblasto en el pollito da como resultado un patrón correctovetas ectópicas , lo que sugiere que el hipoblasto sirve para inhibir la formación de la veta primitiva. [13]
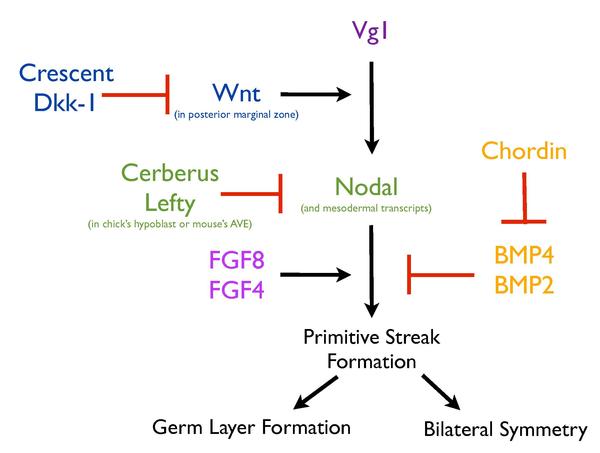
Vg1 y señalización Wnt [ editar ]
De manera similar, la expresión y los injertos de Vg1 (un miembro de la familia TGFB ) de la zona marginal posterior [5]en los pollitos también pueden inducir rayas ectópicas, pero solo dentro de la zona marginal del embrión, [11] lo que indica una característica específica de esta región en su capacidad de inducir la formación de rayas. Varias líneas de evidencia apuntan a la expresión de Wnt como el determinante de esta habilidad. La eliminación de Wnt3 en embriones de ratón da como resultado la ausencia de una formación de estrías, de manera similar al fenotipo de embriones mutantes de B-catenina . [14] Además, mutando el regulador negativo intracelular de señalización de Wnt, Axin , [15] y misexpression de la chica cWnt8C [16] produce múltiples rayas en embriones de ratón. La localización de Wnt y los componentes de su vía, Lef1 y B-catenina, respaldan aún más el papel inductor de rayas en la zona marginal. [11] Además, se expresa como un gradiente que disminuye de posterior a anterior, [11] [12] correspondiente a la capacidad inductora de rayas de la zona marginal. La expresión misex de Vg1 o Wnt1 por sí sola no logró inducir una línea ectópica en el pollito, pero en conjunto su expresión xisex resultó en la formación de líneas ectópicas, lo que confirma que la capacidad inductora de líneas de la zona marginal posterior podría atribuirse a la señalización de Wnt [11] y que Vg1 y Wnt deben cooperar para inducir este proceso. La expresión mixta de Vg1 junto con los antagonistas de Wnt, Crescent o Dkk-1 , previene la formación de rayas ectópicas, [11] lo que demuestra la importancia de la actividad de Wnt en la formación de rayas ectópicas inducidas por Vg1 y, por lo tanto, su implicación en la formación de rayas primitivas normales.
Hipoblasto [ editar ]
Cualquier corte dado del blastodermo es capaz de generar un eje completo [17] hasta el momento de la gastrulación y la formación de vetas primitivas. [13] Esta capacidad de generar una racha a partir del embrión de pollo en etapa previa a la raya [18] indica que debe haber un mecanismo para garantizar que solo se forme una sola raya. El hipoblasto segrega un antagonista de Nodal que evita la formación de estrías ectópicas en el pollito. [13]
Señalización nodal [ editar ]
Nodal , un inductor mesodérmico conocido de la superfamilia TGFB, [18] ha sido implicado en la formación de rayas. Los embriones de ratón mutantes para Nodal no se gastrulan y carecen de la mayoría del mesodermo, [19]pero más que desempeñar un papel en la inducción del mesodermo, Nodal regula la inducción y / o el mantenimiento de la línea primitiva. [19] En presencia de hipoblasto, Nodal no puede inducir rayas ectópicas en el embrión de pollo, mientras que su eliminación, induce la expresión de Nodal, Chordin y Brachyury, [13]sugiriendo que el hipoblasto debe tener un cierto efecto inhibitorio sobre la señalización nodal. De hecho, el antagonista multifuncional de la señalización Nodal, Wnt y BMP, Cerberus (producido en el hipoblasto) y Cerberus-Short (que inhibe solo a Nodal), a través de su efecto sobre la señalización Nodal, inhibe la formación de estrías. [13]Finalmente, el hipoblasto se desplaza anteriormente por el endoblast en movimiento, lo que permite la formación de rayas en el extremo posterior. En el extremo anterior, la presencia del hipoblasto y los antagonistas que secreta, como Cerberus, inhiben la expresión de Nodal y, por lo tanto, restringen la formación de estrías solo en el extremo posterior. [13] De manera similar al hipoblasto en el pollito, el AVE en el ratón segrega dos antagonistas de la señalización nodal, similar a Cerberus, Cerl y Lefty1. [13] [20] En ratón, Cer - / -; Lefty1 - / - mutantes dobles desarrollan múltiples rayas [6] como lo indica la expresión ectópica de Brachyury y pueden ser parcialmente rescatados mediante la eliminación de una copia del gen Nodal. [6] En el mouse, el AVE restringe la formación de rayas a través de las funciones redundantes de Cer1 y Lefty1, que regulan negativamente la señalización nodal. [6] El papel del AVE del ratón para garantizar la formación de una sola racha primitiva se conserva evolutivamente en el hipoblasto del polluelo. [6] [13]
Señalización FGF [ editar ]
Otra vía importante en la formación de modulación de la línea primitiva es FGF , que se cree que trabaja junto con Nodal para regular este proceso. [18] La inhibición de la señalización de FGF a través de la expresión de un receptor negativo dominante, usando un inhibidor del receptor de FGF (SU5402) o el agotamiento de los ligandos de FGF, inhibe la formación de mesodermo [3] y esto a su vez, inhibe la formación de estrías. [4] Además, la formación de estrías ectópicas inducidas por Vg1 requirió señalización de FGF. [18]
Señalización BMP [ editar ]
Finalmente, la señalización BMP también es importante para regular el proceso de formación de estrías en el embrión de pollo. El sitio de formación de rayas se caracteriza por señales bajas de BMP, mientras que el resto del epiblasto muestra altos niveles de activación de BMP. [21] Además, la expresión misex de BMP4 o BMP7 previene la formación de estrías, mientras que el inhibidor de BMP Chordin induce la formación de estrías ectópicas en el pollo, [22] lo que sugiere que la formación de estrías probablemente requiera la inhibición de BMP.
Implicaciones éticas [ editar ]
La raya primitiva es un concepto importante en bioética , donde algunos expertos han argumentado que la experimentación con embriones humanos es permisible, pero solo antes de que se desarrolle la raya primitiva, generalmente alrededor del decimocuarto día de existencia. El desarrollo de la racha primitiva se toma, por tales bioeticistas, para significar la creación de un ser humano único. [23] En algunos países, es ilegal desarrollar un embrión humano durante más de 14 días fuera del cuerpo de una mujer.
No hay comentarios:
Publicar un comentario