TAXONOMÍA DE LA CÉLULA BACTERIANA .-
Diferencias entre procarióntes y eucariontes
La célula de los procarióntes, en contraste con la célula eucariónte, no es compartamentalizada. No están presentes, membranas nucleares, ni mitocondria, ni retículo endoplásmico, ni aparato de Golgi, ni fagosomas o lisosomas. (Figura 1b, 2 y 3). Los procarióntes generalmente poseen un sólo cromosoma circular. Debido a que no hay membrana nuclear, el cromosoma está unido a un sitio específico en la membrana celular - el mesosoma. Los ribosomas procarióntes son 70S (S representa la unidad de Svedberg, una medida de tamaño), mientras que ribosomas eucariontes son más grandes (80S). Las subunidades ribosomales procariontes son 30S y 50S (en los eucariontes son más grandes). La subunidad ribosomal 30S contiene RNA 16S, mientras que la subunidad ribosomal 50S contiene 23S y 5S. El RNA ribosomal es mayor en los eucariontes (por ejemplo. rRNA 18S contra 16S). Las membranas bacterianas generalmente no contienen esteroles (por ej., colesterol).
Estructuras Bacterianas
A pesar de que comparadas con los eucariontes, estas carecen de complejidad, se pueden definir estructuras eubacteriales, aunque no todas las bacterias poseen cada uno de estos componentes.
Plásmidos
Estas son moléculas de DNA extracromosomal, presentes generalmente en múltiples copias, que a menudo codifican para factores de patogenicidad y factores de resistencia a antibióticos. Algunas formas también están relacionadas con la replicación bacteriana.
La envoltura celular
Las bacterias se pueden dividir en dos grupos sobre las bases de su tinción de Gram. Las bacterias gram positivas se quedan teñidas con cristal violeta después de lavar y las gram negativas no. Todas las bacterias tienen una membrana celular donde ocurre la fosforilación oxidativa (ya que no tienen mitocondrias). Al exterior de la membrana celular, está la pared celular, la cual es rígida y protege a la célula de la lisis celular. En las bacterias gram positivas, la capa de peptidoglicano de su pared celular es una capa mucho más gruesa que en las bacterias gram negativas. Las bacterias gram negativas tienen una membrana externa adicional. La membrana externa es la barrera más importante de permeabilidad en las bacterias gram negativas. El espacio entre las membranas interna y externa se conoce como espacio periplásmico. En el espacio periplásmico las bacterias Gram negativas almacenan enzimas degradativas. Las bacterias Gram positivas carecen de espacio periplásmico; en su lugar secretan exo-enzimas y realizan digestión extracelular. Esta digestión es necesaria ya que moléculas mas bien grandes no pueden pasar fácilmente a través de la membrana externa (si está presente) o la membrana celular.
|
| |
Formas Bacterianas libres de pared
Cuando las bacterias se tratan 1) con enzimas líticas que degradan la pared celular, por ejemplo: lisozima o 2) con antibióticos que interfieren con la biosíntesis de peptidoglicanos, se producen bacterias libres de pared celular. Generalmente estos tratamientos generan organismos no viables, pero que son útiles en la experimentación in vitro. A las bacterias libres de pared que no pueden replicar se les llama esferoplastos (cuando la membrana externa aún esta presente) o protoplastos (si la membrana externa ya no está presente). Ocasionalmente estos tratamientos generan formas de bacterias libres de pared que pueden replicar (las formas L).
|
 Figura 4. Bacilo E. coli –procarionte con múltiples flagelos (SEM x26,730) © la Microscopia, S.a. Utilizado con permisode Dennis Kunkel Figura 4. Bacilo E. coli –procarionte con múltiples flagelos (SEM x26,730) © la Microscopia, S.a. Utilizado con permisode Dennis Kunkel
|
Flagelos
Algunas especies bacterianas son móviles y poseen organelos de locomoción – los flagelos (Figura 4), aquellas que los tienen son capaces de sentir su ambiente y responder a nutrientes químicos o materiales tóxicos y mueven hacia ellos o se alejan de ellos (por el fenómeno llamado quimiotáxis). Los flagelos están embebidos en la membrana celular, y se extienden a través de la envoltura celular proyectándose como largas cadenas. Los flagelos consisten de un número de proteínas que incluyen la flagelina. Los flagelos mueven a la célula mediante rotación por una acción tipo propela. Los filamentos axiales en las espiroquetas tienen una función similar a los flagelos. Las proteínas de unión en el espacio periplásmico o en la membrana de célula se unen a fuentes de alimento (tal como azúcares y aminoácidos) causando metilación de otras proteínas celulares de membrana, las cuales a su vez afectan el movimiento de la célula por el flagelo. Las permeasas son proteínas que transportan estos alimentos a través de la membrana celular. Las fuentes de carbono y energía pueden ser almacenadas cuando sea necesario en “gránulos citoplásmicos” los cuales consisten de glicógeno, polihidroxibutirato o polifosfato.
|
 Figura 5. E. coli con fimbrias (TEM x17,250) © la Microscopia de Dennis Kunkel, S.a. Utilizado con el permiso Figura 5. E. coli con fimbrias (TEM x17,250) © la Microscopia de Dennis Kunkel, S.a. Utilizado con el permiso |
Pili (sinónimo: fimbrias)
Los tipos de pili (cuando están presentes) varían entre las especies. Los pili son proyecciones tipo fibroso de las células (Figura 5). Algunos están relacionados con la conjugación sexual y otros permiten la adherencia de la bacteria a las superficies epiteliales del huésped que infecta.
|
 Figura 6. Bacterias con forma de bacilo formadoras de cápsula. La cápsula está compuesta de polisacáridos y poliproteínas. Las cápsulas tienen un papel en la adherencia, virulencia, protección, aseguramiento de nutrientes y reconocimiento célula-célula. Las cápsulas varían en grosor y pueden fácilmente representar 2 veces el volumen del organismo. En una tinción de cápsula, el fondo se tiñe de azul grisáceo y las células se tiñen en rojo. La cápsula no esta teñida y aparece como una halo alrededor de la célula. © Judy Bowen Figura 6. Bacterias con forma de bacilo formadoras de cápsula. La cápsula está compuesta de polisacáridos y poliproteínas. Las cápsulas tienen un papel en la adherencia, virulencia, protección, aseguramiento de nutrientes y reconocimiento célula-célula. Las cápsulas varían en grosor y pueden fácilmente representar 2 veces el volumen del organismo. En una tinción de cápsula, el fondo se tiñe de azul grisáceo y las células se tiñen en rojo. La cápsula no esta teñida y aparece como una halo alrededor de la célula. © Judy Bowen
Buckman Laboratories International, Inc.
Memphis, Tennessee, USA and The MicrobeLibrary
|
Cápsulas y laminas viscosas (Figura 6)
Se trata de estructuras que rodean el exterior de la envoltura celular. Cuándo se ven más definidas nos referimos a ellas como la cápsula. Cuando se observan menos definidas, nos referimos a ellas como lámina viscosa o glicocalix. Estas estructuras usualmente consisten de polisacáridos; sin embargo, en ciertos bacilos, están compuestas de un polipéptido (el ácido poliglutámico). Estas estructuras no son esenciales a la viabilidad celular y en algunas especies habrá cepas que producirán cápsula, mientras que otras no. Las cápsulas de las bacterias inhiben la ingestión y muerte producida por los fagocitos. Durante el cultivo in vitro la síntesis de la cápsula a menudo se pierde.
Endosporas (esporas)
Estas son formas latentes de células bacterianas producidas por ciertas bacterias en condiciones de ayuno (Figura 7); las formas de crecimiento activo de la célula se llaman células vegetativas. La espora es resistente a condiciones adversas (incluyendo temperaturas altas y solventes orgánicos). El citoplasma de la espora es deshidratado y contiene dipicolinato de calcio (ácido dipicolínico) (Figura 8) el cual está relacionado con la resistencia de la espora al calor extremo. Las esporas se encuentran comúnmente en los géneros Bacillus yClostridium.
|
| |  Figure 7 A. Espora de Bacillus cereus (verde) y las células que no forman esporas (rosa) © Ralph Van Dyke Jr. The Des Moines University Osteopathic Medical Center, Des Moines, Iowa USA and The MicrobeLibrary Figure 7 A. Espora de Bacillus cereus (verde) y las células que no forman esporas (rosa) © Ralph Van Dyke Jr. The Des Moines University Osteopathic Medical Center, Des Moines, Iowa USA and The MicrobeLibrary | 
Figura 7B. larvae esporulación el cultivo se muestra de forma similar a las de otros formadores de esporas. La célula vegetativa en forma de bastón tiene una gruesa de capa de peptidoglicano. © Donald Stahly, la University of Iowa, Iowa City, and The MicrobeLibrary | 
Figura 7C. Una espora inmadura se muestra rodeada por su célula madre (esporangio). Una copia del DNA bacteriano esta contenida dentro de la espora en desarrollo. La cubierta externa de la espora aparece más delgada y menos electrodensa que en la célula madura. © Donald Stahly, la University of Iowa, Iowa City, and The MicrobeLibrary | 
Figura 7D. La cubierta gruesa de la espora indica que la diferenciación de la endospora ya es completa, aunque la endospora permanece aún dentro del esporangio. Finalmente, la endospora se libera del esporangio. La cubierta interna consiste en un máximo de siete capas distintas llamadas “lamellae”. © Donald Stahly, la University of Iowa, Iowa City, and The MicrobeLibrary |
|  Figura 8. Acido Dipicolínico Figura 8. Acido Dipicolínico | |
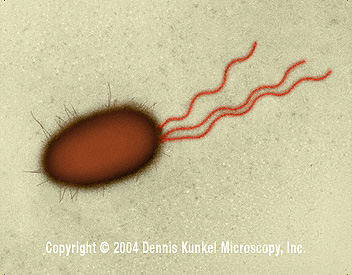 Figura 4. Bacilo E. coli –procarionte con múltiples flagelos (SEM x26,730) © la Microscopia, S.a. Utilizado con permisode Dennis Kunkel
Figura 4. Bacilo E. coli –procarionte con múltiples flagelos (SEM x26,730) © la Microscopia, S.a. Utilizado con permisode Dennis Kunkel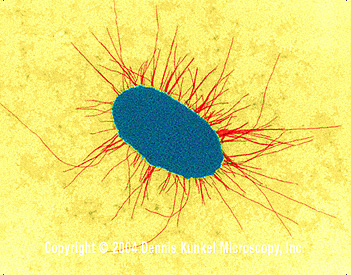 Figura 5. E. coli con fimbrias (TEM x17,250) © la Microscopia de Dennis Kunkel, S.a. Utilizado con el permiso
Figura 5. E. coli con fimbrias (TEM x17,250) © la Microscopia de Dennis Kunkel, S.a. Utilizado con el permiso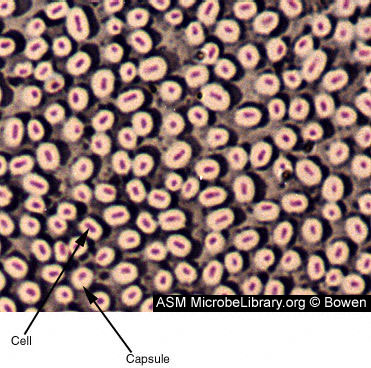 Figura 6. Bacterias con forma de bacilo formadoras de cápsula. La cápsula está compuesta de polisacáridos y poliproteínas. Las cápsulas tienen un papel en la adherencia, virulencia, protección, aseguramiento de nutrientes y reconocimiento célula-célula. Las cápsulas varían en grosor y pueden fácilmente representar 2 veces el volumen del organismo. En una tinción de cápsula, el fondo se tiñe de azul grisáceo y las células se tiñen en rojo. La cápsula no esta teñida y aparece como una halo alrededor de la célula. © Judy Bowen
Figura 6. Bacterias con forma de bacilo formadoras de cápsula. La cápsula está compuesta de polisacáridos y poliproteínas. Las cápsulas tienen un papel en la adherencia, virulencia, protección, aseguramiento de nutrientes y reconocimiento célula-célula. Las cápsulas varían en grosor y pueden fácilmente representar 2 veces el volumen del organismo. En una tinción de cápsula, el fondo se tiñe de azul grisáceo y las células se tiñen en rojo. La cápsula no esta teñida y aparece como una halo alrededor de la célula. © Judy Bowen Figure 7 A. Espora de Bacillus cereus (verde) y las células que no forman esporas (rosa) © Ralph Van Dyke Jr. The Des Moines University Osteopathic Medical Center, Des Moines, Iowa USA and The MicrobeLibrary
Figure 7 A. Espora de Bacillus cereus (verde) y las células que no forman esporas (rosa) © Ralph Van Dyke Jr. The Des Moines University Osteopathic Medical Center, Des Moines, Iowa USA and The MicrobeLibrary 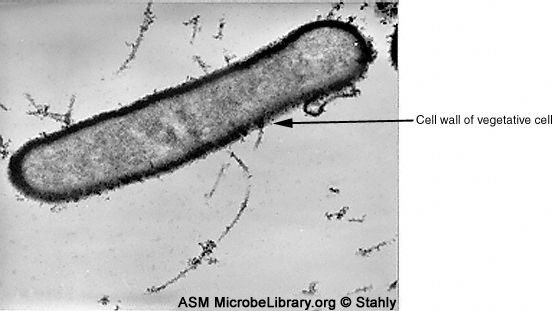

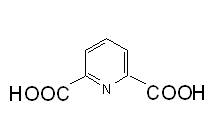 Figura 8. Acido Dipicolínico
Figura 8. Acido Dipicolínico
No hay comentarios:
Publicar un comentario