acanthocephala
Vamos a seguir el esquema de clasificación utilizado por Roberts y Janovy. La clasificación de acanthocephala es difícil debido a que comparten tan pocas características con otros grupos. Ellos se han asociado con diversas nematodos y rotíferos en el tiempo y el trabajo reciente sugiere que los rotíferos son sus parientes más cercanos. Cuando se encuentre con términos que no son familiares se refieren a su glosario para las definiciones.
Características generales
Acanthocephala se reconoció por primera vez en el siglo 17, pero no fue hasta el siglo 20 que se reconocen como un filo independiente. Raramente se encuentran en los seres humanos y los animales domésticos, por lo que la mayoría de los médicos no están familiarizados con el grupo. La excepción a esto es Macracanthorhynchus hirudinaceus , el cual es un parásito cosmopolita de cerdos. Todos los miembros del filo son parásitas. El ciclo de vida por lo general implica un huésped definitivo de vertebrados y artrópodos huésped intermediario.
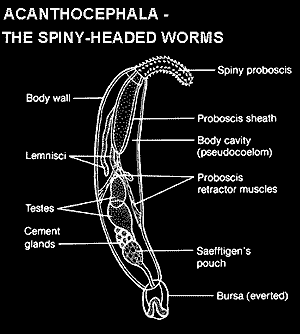
Acanthocephala son poco más que bolsas de gónadas. En ambos sexos, se reduce el sistema nervioso, sistema muscular, sistema excretor, y el sistema circulatorio. El aparato digestivo está completamente ausente. Los gusanos son pseudocoelomate con unas ventosas trompa en el extremo anterior. La figura 31.1 ilustra la estructura general del cuerpo de un acantocéfalo. Características destacadas son la trompa, el cuello y el tronco . El, trompa hueca llena de líquido está cubierta con ganchos y su forma varía según las especies de cilíndrica a esférica (Figura 31.2). El tamaño, la forma y número de ganchos son caracteres taxonómicos importantes. Músculos Proboscis retractor están unidos al vértice interior de la trompa y se extienden la longitud de la trompa y el cuello antes de insertar en un saco muscular conocido como el receptáculo proboscis (Figuras 31.1 y 31.3). La contracción de los músculos retractores trompa invagina la trompa y los ganchos son entonces completamente en el interior. Evaginación de la trompa se efectúa por la contracción del recipiente trompa, lo que obliga a la trompa a cabo mediante un sistema hidráulico. El ganglio cerebral o cerebro se encuentra dentro del receptáculo trompa.
Presoma es el nombre colectivo de la trompa y su receptáculo. El cuello es suave, unspined y entre el extremo posterior de la trompa y el tronco. Algunas especies poseen músculos retractores del cuelloque trabajan con los músculos retractores trompa de retirar todo el extremo anterior en el maletero. Un pozo sensorial se encuentra en cada lado del cuello en algunas especies, mientras que muchas especies tienen un par de pozos similares en la punta de la trompa.
Metasoma es otro nombre para el tronco. Está cubierta por un tegumento (como son la trompa y el cuello) y tiene capas internas musculares. Algunas especies tienen espinas incrustadas en el tegumento que aparecen para ayudar a mantener un estrecho contacto con la mucosa intestinal del huésped. El sistema reproductor se encuentra en el maletero. El tronco absorbe los nutrientes de acogida y ayuda a distribuir dentro del gusano. Gusanos vivos tienen un tronco arrugado, bilateralmente aplanado que se convertirá en turgentes cuando se ponen en una solución hipotónica. Este fenómeno se utiliza en la preparación de los gusanos para el estudio, ya que generalmente obliga a la trompa de evaginar, que es importante en la identificación de la especie.
Wall cuerpo
La pared del cuerpo es sincitial con numerosos núcleos y una compleja serie de canales internos, interconectadas llamadas al sistema lacunar (Figuras 31.4, 31.5 y 31.6). Los núcleos son grandes y pocos en número en algunas especies, mientras que en otros se fragmentan durante el desarrollo y están ampliamente distribuidos en la pared del cuerpo. Las especies con pocos núcleos de exposiciones eutelia , o la constancia nuclear. Esto, también, puede servir como un carácter taxonómico.
tegumento
Hay varias capas distintas al tegumento (Figura 31.4). De afuera hacia adentro, que son:
- la capa de superficie
- la zona rayada
- la zona de fieltro de fibra
- la zona radial de la fibra
- la membrana basal .
La zona rayada es de 4-6 um de espesor y es perforada por criptas que son 2-4 um profunda y abierta a la superficie por los poros. Las criptas dan un aspecto de la zona rayada bajo el microscopio óptico, de ahí el nombre. Las criptas aumentan en gran medida el área superficial. Los cuellos de las criptas tienen un tamiz molecular filamentoso a través del cual las partículas de menos de 8,5 nm puede pasar. Estas partículas pueden someterse a pinocitosis por la membrana cripta, a pesar de la importancia de la pinocitosis en la adquisición de nutrientes todavía no se comprende totalmente. Numerosas gotas de lípidos, mitocondrias, complejos de Golgi, los lisosomas y se encuentran en las regiones más profundas de la zona.
La zona de fieltro de fibra está al lado y contiene mitocondrias, numerosas partículas de glucógeno, vesículas, y de vez en cuando lisosomas y las gotas de lípidos. Justo dentro de la zona de fieltro de fibra es la zona radial de la fibra que constituye alrededor del 80% del espesor de la pared del cuerpo. Dentro de la zona radial de la fibra son grandes haces de filamentos que pasan radialmente a través del citoplasma, núcleos de la pared del cuerpo, y grandes gotas de lípidos. partículas de glucógeno, complejos de Golgi, lisosomas, mitocondrias y también se encuentran en esta zona. El citoplasma perinuclear contiene retículo endoplasmático rugoso. Los canales lacunares pasan a través de la zona radial de la fibra.
La pared proboscis es similar en estructura a la pared del cuerpo, pero tiene un menor número de criptas, una zona radial más delgada, y no hay zona de fieltro de fibra.
Sistema lagunar y los músculos
La función del sistema lagunar ha sido poco clara, pero la investigación realizada por el Dr. mis amigos Tommy Dunagan y el fallecido Dr. Donald Miller ha ayudado a dilucidar la función y la asociación del sistema con la capa de músculo circular.
Hay dos partes no conectadas al sistema lagunar, (1) el sistema presomal en la trompa y el cuello, y (2) el sistema metasomal asociados con la pared del cuerpo. Los canales en el sistema presomal se ejecutan en un par de lemniscos , que son extensiones de la zona radial de la fibra que crece desde la base del cuello hasta la pseudoceloma. El canal central de cada lemniscus es continuo con el sistema lagunar presomal. La función lemniscal es desconocida, pero se ha sugerido que contribuyen a la hidráulica del mecanismo de trompa.
El sistema metasomal lacunar es una compleja red de canales interconectados. La disposición y la ubicación de la lacuni son específicos para cada especie y servir como caracteres taxonómicos útiles. La mayoría de las especies tienen un par de canales principales longitudinales o bien laterales o dorsal y ventral en el arreglo. Los canales principales están conectados por una serie de canales transversales (Figura 31.5). Algunas especies tienen un par de canales longitudinales mediales conectados a la a la otra dorsal y ventral canales por canales radiales cortos unidos a canales de anillo circulares (figura 31.5).Los canales mediales están en el lado pseudocoel de la pared del cuerpo y los canales radiales pasan a través de los músculos para conectarse a los canales anulares.
Los músculos del cuerpo de pared incluyen una capa longitudinal interior rodeado por una capa circular exterior (figura 31.6). Los músculos tienen una estructura hueca inusual con numerosas anastomosis de interconexión. Las luces de los músculos son continuas con el sistema lagunar lo que sugiere que la circulación del líquido lacunar podría servir para llevar los nutrientes a eliminar los desechos y de los músculos. La contracción de los músculos longitudinales obligaría fluido en los músculos circulares y viceversa. Por lo tanto, el pensamiento actual es que el sistema lacunar sirve como un sistema de transporte de fluido en combinación con un esqueleto hidrostático.
Acantocéfalo músculos son diferentes a los músculos en otros organismos, ya que tienen bajos potenciales de membrana, son eléctricamente inexcitable, y exhiben conducción lenta. La despolarización de los músculos es espontánea y rítmica. La acetilcolina parece estimular los músculos, pero el control de la contracción nerviosa no ha sido bien demostrado. Si el control de sistema nervioso se produce se cree que hacerlo a través del sistema de rete , que es una red muy ramificada, de anastomosis de los túbulos de pared delgada que se encuentran en la superficie medial de los músculos longitudinales o entre las capas longitudinales y circulares. El sistema rete se puede modificar las células musculares.
Sistema nervioso
El sistema nervioso es reducida y muy simple en el acanthocephala. El ganglio cerebral se encuentra en el receptáculo proboscis y tiene relativamente pocos nervios que salen de ella. El nervio trompa anterior y posterior de los nervios laterales son más grandes que los nervios se ramifican desde el ganglio. Los nervios suministran los dos órganos de los sentidos laterales y apicales órgano de los sentidos, si está presente. Un grande, multinucleadas célula de apoyo se encuentra ventral y ligeramente por delante del ganglio cerebral. Procesos conducen desde esta celda para los órganos de los sentidos, pero no son los nervios y su función es desconocida. Ellos pueden tener una función secretora que puede ayudar a explicar la respuesta inflamatoria del huésped a la trompa del gusano.
Sistema Excretor
La mayoría de acanthocephala utilizan difusión a través de la pared del cuerpo para efectuar la excreción. Sin embargo, los miembros de la familia Oligoacanthorhynchida poseen dos órganos excretores protonephridial . Estos órganos tienen numerosas bombillas de llama anucleadas con mechones de flagelos que pueden o no estar encapsulados. En las mujeres estos órganos están asociadas a la campana uterina (véase más adelante) y desembocan en el útero; en los hombres que están unidos a los conductos deferentes y vacío a través de él.
osmoregulatorias capacidades están limitadas en el Acanthocephala. En una solución hipotónica y se hinchan en soluciones hipertónicas que se vuelven flácidas. La presión osmótica de fluidos pseudocoelomic está cerca o ligeramente superior a la de los contenidos intestinales del huésped.
Reproducción
Acantocéfalos tienen sexos separados y las hembras son generalmente más grandes que los machos (Figura 31.3). Sacos de ligamentos están unidos al extremo posterior de la trompa y se extienden hasta cerca de los poros genitales distales de ambos sexos. Gónadas y órganos accesorios están contenidos dentro de estos sacos. En algunas especies, los sacos de ligamentos son permanentes; en otros se descomponen como el gusano madura.
Sistema de Mujer
Acantocéfalos tienen un ovario muy singular que fragmenta en bolas de ovario , a menudo antes de la maduración. Las bolas de ovario consisten en ovogonia flotando libremente en el saco del ligamento.Las bolas aumentan ligeramente de tamaño antes de la inseminación. A muscular de campana uterina está unido al extremo posterior del saco ligamento (Figura 31.8). Este órganos ordena los huevos maduros de los huevos inmaduros, permitiendo que los maduros para pasar en el útero y la vagina y hacia fuera del poro genital. Los huevos inmaduros se devuelven a la salida del ligamento.
Tras la cópula, los espermatozoides migran desde la vagina, a través del útero y el útero de campana en el saco del ligamento. Los ovocitos en las bolas de ovario se fertilizan y después de las primeras divisiones se separan de las bolas de ovario que exponen los ovocitos subyacentes. La fertilización se tambaleó cuando los ovocitos son más exteriores etapas primera y varios fertilizados de la embriogénesis temprana se pueden encontrar en una sola hembra. Los embriones en desarrollo flotan libremente en el líquido pseudocoelomic sometidos a un mayor desarrollo como embriones sin cáscara . Embriones totalmente desarrollados son llamados acanthors sin cáscara .
Los acanthors sin cáscara son empujados en la campana uterina por la acción peristáltica y pueden seguir una de dos rutas. Si ellos no están completamente desarrollados pasarán a través de ranuras en la campana y volver a la pseudoceloma. embriones maduros son demasiado grandes para pasar a través de las rendijas y continuarán a través del útero, la vagina y, finalmente, el poro genital en el intestino del huésped. Este mecanismo es bastante eficiente y la investigación sugiere que no hay embriones inmaduros son liberados en el útero.
Sistema de sexo masculino
En la mayoría de las especies se producen dos testículos en los machos y la ubicación y el tamaño son bastante consistentes (Figura 31.3). Espermatozoides maduros pasar de cada testículo en un efferens conductos que conducen a un común conductos deferentes o pene. Los espermatozoides son sin cabeza, hilos delgados. Varios órganos accesorios están presentes en el sistema masculino incluyendo lasglándulas de cemento , que son órganos sincitiales con uno o más núcleos gigantes o varios fragmentos nucleares. Cuando más de una glándula está presente están unidos por conexiones esbeltas. Las glándulas de cemento secretan una proteína bronceada llamado cemento copulador , que en algunas especies se almacena en un depósito de cemento . El cemento es secretada después de la cópula y forma un tapón de copulador o tapón de cemento que bloquea la vagina, evitando la fertilización por otros machos. La tapa finalmente se desintegrará, por lo que acanthors sin cáscara completamente desarrolladas pueden ser liberados.
La bursa copulador (Figura 31.7) es una extensión especializada, en forma de campana de la pared del cuerpo que permanece invaginado en el extremo posterior del cuerpo, excepto durante la cópula. Labolsa de Saefftigen es un saco muscular unido a la base de la bolsa que los contratos y las fuerzas del fluido en la bolsa de Evert que por la presión hidrostática. Línea papilas sensoriales de la bursa y cuando hace contacto con una hembra, que a su Corchetes por la contracción muscular y la transferencia de esperma se produce a través del pene.
Adquisición de Nutrientes
La absorción de nutrientes y el metabolismo de una sola especie ( moniliformis moniliformis ) ha sido bien estudiados, aunque algunos trabajos se ha hecho en otras especies. Por lo tanto, las generalizaciones acerca de la aplicabilidad de los resultados del estudio es problemático. La mayor parte de la información a continuación se basa en las pocas especies que se han estudiado.
Consumo
Todas las moléculas de nutrientes deben ser tomadas a través de la pared del cuerpo, al igual que en la Cestoda.
Los triglicéridos son absorbidos por el presoma y, finalmente, se acumulan en el lemniscos. Los aminoácidos son absorbidos, en parte, por los sistemas de transporte de membrana estereoespecíficos. La superficie de M. moniliformis contiene peptidasas que pueden escindir varios dipéptidos para permitir la absorción de los aminoácidos resultantes por el gusano. En algunas especies de lisina se absorbe a través de la tegumento metasomal y se acumula en los núcleos y el cinturón muscular exterior. La timidina se absorbe y se incorpora en el ADN en las regiones perilacunar y en los núcleos de las bolas de ovario y testículos. Timidina radiactiva no se encuentra en los núcleos de la pared del cuerpo y se supone que el ADN sintetizado en la pared es mitocondrial.
M. moniliformis absorbe la totalidad de sus hidratos de carbono de su anfitrión. El gusano puede absorber manosa, glucosa, fructosa, galactosa, y varios análogos de glucosa. El locus de la glucosa sirve como el sitio para la absorción de la glucosa, mientras que los otros hidratos de carbono se absorben en el denominado sitio de fructosa . La maltosa y glucosa-6-fosfato se hidrolizan en glucosa por las enzimas antes de ser absorbido. A diferencia de la mayoría de los sistemas de transporte de la glucosa, el sistema de M. moniliformis no está acoplado a la co-transporte de sodio.
Metabolismo energético
Metabolismo energético acanthocephalan está adaptado para anaerobiosis facultativa. M. moniliformis puede fermentar las hexosas que son absorbidos, pero el ciclo de Krebs no parece estar presente la M.moniliformis o Macracanthorhynchus hirudinaceus . Sin embargo, Echinorhynchus Gadi hace evidenciar el ciclo de Krebs. Los productos finales de la glucólisis en M. moniliformis son etanol y lactato, en lugar de succinato.
Lípidos endógenos no se metabolizan durante la incubación in vitro de M. moniliformis que sugieren que los lípidos no se utilizan como fuente de energía. Las enzimas requeridas para la beta-oxidación de los lípidos muestran bajos niveles de actividad o están ausentes por completo.
Poco se sabe sobre el transporte de electrones en acantocéfalos.
Los ciclos de desarrollo y de la vida (Figuras 31.9, 31.10, 31.11)
Ciclos de vida
La figura 31.9 ilustra un ciclo de vida típico de un acantocéfalo. Los artrópodos constituyen el huésped intermediario y el insecto, crustáceos, miriápodos o debe comer un "huevo" (en realidad un acanthor cáscara) que se publicó con las heces del huésped definitivo de los vertebrados. Una vez dentro del huésped intermediario, el parásito se desarrolla a través de varias etapas de la última de las cuales es infecciosa para el huésped definitivo. El huésped definitivo consume un huésped intermediario infectada a continuar con el ciclo de la vida. Un anfitrión paraténico es un vertebrado hospedador definitivo no aptos en la que el parásito no madurará, sino que enquistarse en las vísceras peritoneal. Si el anfitrión paraténico es comido por el huésped definitivo adecuado, se completará el ciclo de vida. Si no, es un callejón sin salida para el parásito. Un anfitrión paraténico puede cerrar la brecha entre microcrustacea y grandes peces se alimentan de peces. Los peces pequeños en la Figura 31.9 sirve como un ejemplo de un anfitrión paraténico.
Vamos a examinar el ciclo de vida de dirus Acanthocephalus a través de material en vivo en algún momento de este semestre.
Desarrollo
Segmentación temprana es espiral y ligeramente distorsionada por la forma de huso de la cáscara del huevo. Entre el estado de 4 celdas y la etapa de 34 células, límites de las celdas desaparecen y todo el embrión se convierte en sincitial. Los núcleos migran hacia el interior durante la gastrulación. Los núcleos se hacen más pequeños a medida que continúan dividiéndose y, finalmente, forman un núcleo denso de pequeños núcleos llama la masa interna . Todos los órganos internos del gusano desarrollan a partir de estos núcleos. Dependiendo de la especie, el tegumento desarrolla a partir de los núcleos no condensados restantes en la zona periférica; o a partir de un núcleo que se separa de la masa interna, o de las contribuciones de ambos.
El acanthor está totalmente embrionados y infeccioso respecto a la huésped intermediario de artrópodos. Es un organismo alargado frecuencia que posee seis u ocho ganchos bladelike en el extremo anterior.Estos ganchos ayudar en la penetración del intestino del huésped intermediario. En algunas especies, los ganchos son reemplazadas por pequeñas espinas. Juntos, los ganchos o espinas y sus músculos se llaman el órgano aclid o rostellum . Acanthors no desarrollará más hasta que es ingerido por un huésped intermediario adecuado. Acanthors pueden permanecer viables durante meses en condiciones ambientales normales. El acanthor penetrará en el intestino del huésped a cualquiera de entrar de lleno en la hemocoel o mentir justo debajo de la serosa. El acanthor comenzará a absorber los nutrientes de acogida y desarrollar aún más en la próxima etapa larval del Acanthella . Durante la etapa de Acanthella, los sistemas de órganos se desarrollan a partir de la masa nuclear central y los núcleos hipodermis.
La etapa de desarrollo final es la cistacanto , que es infectivo para el huésped definitivo. No se observa evolución ocurrirá hasta que el cistacanto es ingerido por un huésped definitivo adecuado. Los extremos anterior y posterior de la cistacanto se invaginan y todo el gusano está encerrado en una vaina hialina. Sólo una pequeña fracción de la gran cantidad de huevos producidos jamás llegar a la etapa cistacanto y algunos de éstos se ingiere por un huésped definitivo.
Efectos de acantocéfalos en sus anfitriones
Una de las características fascinantes de acanthocephala es su influencia en el comportamiento, morfología, y otras características de sus huéspedes intermediarios que tienden a aumentar la probabilidad de transmisión a los huéspedes definitivos. Las cucarachas ( Periplaneta americana y Blatella germanica ) infectados con M. moniliformis viajan distancias más cortas, se mueven más lentamente, y pasar más tiempo en las superficies horizontales de hacer los controles no infectados. Isópodos acuáticos ( Caecidotea intermedius ) infectados con dirus Acanthocephalus son hiperactivos, pasar más tiempo en la parte superior de la hojarasca, y puede ser notablemente más ligero en la pigmentación que son isópodos no infectadas.
Anfípodos ( Gammarus lacustris ) infectados con Polymorphus paradoxus cambian su respuesta fototácticas de negativo a positivo y el anfípodos infectados permanecer en la superficie del agua en lugar de buceo en respuesta a las perturbaciones. Incluso pueden aferrarse a la vegetación flotante. Estas modificaciones del comportamiento mejoran la transmisión de los gusanos a sus huéspedes definitivos (rata almizclera y patos alimentan en la superficie).
Acantocéfalos pueden causar graves daños a los intestinos de sus anfitriones. perforación completa del intestino (Figura 31.12) puede ocurrir y esto es a menudo fatal. La infección bacteriana secundaria puede dar lugar a una peritonitis, hemorragia, pericarditis, miocarditis, arteritis, cholangiolitis, y otros problemas.
A pesar de que puede producirse una perforación, por lo general hay poca respuesta inflamatoria a la penetración de la trompa a pesar de encapsulación puede ocurrir (Figura 31.13). Al menos una especie (M. hirudinaceus ) hace producir antígenos que provocan una respuesta inflamatoria intensa, pero esta especie parece ser la excepción.



No hay comentarios:
Publicar un comentario