 | |||
| |||
| Los nombres | |||
|---|---|---|---|
| Nombre IUPAC
9 H -purin-6-amina
| |||
| Otros nombres
6-aminopurina
| |||
| Identificadores | |||
| CHEBI | |||
| CHEMBL | |||
| ChemSpider | |||
| DrugBank | |||
| ECHA InfoCard | 100.000.724 | ||
| Número de EC | 200-796-1 | ||
| KEGG | |||
PubChem CID
| |||
| Numero de RTECS | AU6125000 | ||
| UNII | |||
| Propiedades | |||
| C 5 H 5 N 5 | |||
| Masa molar | 135.13 g / mol | ||
| Apariencia | Blanco a amarillo claro, cristalino. | ||
| Densidad | 1,6 g / cm 3 (calculado) | ||
| Punto de fusion | 360 to 365 °C (680 to 689 °F; 633 to 638 K) decomposes | ||
| 0.103 g/100 mL | |||
| Solubility | negligible in ethanol | ||
| Acidity (pKa) | 4.15 (secondary), 9.80 (primary)[1] | ||
| Thermochemistry | |||
Heat capacity (C)
| 147.0 J/(K·mol) | ||
Std enthalpy of
formation(ΔfH⦵298) | 96.9 kJ/mol | ||
| Hazards | |||
| Safety data sheet | MSDS | ||
| Lethal dose or concentration (LD, LC): | |||
LD50 (median dose)
| 227 mg/kg (rat, oral) | ||
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa).
| |||
| Infobox references | |||


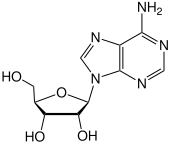
Adenina / æ d ɪ n ɪ n / ( A , Ade ) es una nucleobase (a purinaderivado). Es una de las cuatro nucleobases en el ácido nucleicodel ADN que están representadas por las letras G – C – A – T. Los otros tres son guanina , citosina y timina . Sus derivados tienen una variedad de funciones en la bioquímica que incluye la respiración celular , tanto en forma de trifosfato de adenosina rico en energía (ATP) como encofactores nicotinamida adenina dinucleótido (NAD) y flavina adenina dinucleótido (FAD). También tiene funciones en la síntesis de proteínas y como un componente químico del ADN y el ARN . [2] La forma de la adenina es complementaria a la timina en el ADN o al uracilo en el ARN .
La imagen adyacente muestra adenina pura, como una molécula independiente. Cuando se conecta al ADN, se forma un enlace covalente entre el azúcar desoxirribosa y el nitrógeno de la parte inferior izquierda, eliminando así el hidrógeno. La estructura restante se llama un residuo de adenina , como parte de una molécula más grande. La adenosina es adenina reaccionada con ribosa como se usa en ARN y ATP; desoxiadenosina, adenina unida a la desoxirribosa , como se usa para formar el ADN.
Estructura [ editar ]

La adenina forma varios tautómeros , compuestos que pueden interconvertirse rápidamente y que a menudo se consideran equivalentes. Sin embargo, en condiciones aisladas, es decir, en una matriz de gas inerte y en la fase gaseosa, se encuentra principalmente el tautómero 9H-adenina. [3] [4]
Biosíntesis [ editar ]
El metabolismo de la purina implica la formación de adenina y guanina . Tanto la adenina como la guanina derivan del nucleótido inosina monofosfato (IMP), que a su vez se sintetiza a partir de un fosfato de ribosa preexistente a través de una vía compleja que utiliza átomos de los aminoácidos glicina , glutamina y ácido aspártico , así como la coenzima. tetrahidrofolato .
Tanto la adenina como la guanina derivan del nucleótido inosina monofosfato (IMP), una clase de estructuras moleculares que se han convertido en ácidos nucleicos, la mitad de la dualidad de las funciones de los ácidos nucleicos, la otra mitad son los elementos atómico-moleculares que bioquímicamente y la biología molecular / celular forma el proceso dinámico y caótico de la vida de formas de vida basadas en carbono orgánico. IMP es la estructura molecular "plantilla" para los nucleótidos, por lo tanto la estructura molecular de los ácidos nucleicos existentes.
Función [ editar ]
La adenina es una de las dos nucleobases de purina (la otra es la guanina ) utilizada en la formación de nucleótidos de los ácidos nucleicos . En el ADN, la adenina se une a la timina a través de dos enlaces de hidrógeno para ayudar a estabilizar las estructuras de ácido nucleico. En el ARN, que se usa para la síntesis de proteínas , la adenina se une al uracilo .
 |  |  |  |
| AT-Base-par (ADN) | AU-Base-par (ARN) | AD-base-par (ARN) | A-Ψ-Base-par (ARN) |
La adenina forma adenosina , un nucleósido , cuando se une a la ribosa , y desoxiadenosina cuando se une a la desoxirribosa . Forma adenosina trifosfato (ATP), un nucleósido trifosfato , cuando se agregan tres grupos fosfatoa la adenosina. El trifosfato de adenosina se usa en el metabolismo celular como uno de los métodos básicos para transferir energía química entre reacciones químicas .

Adenosina, a Desoxiadenosina, dA


Historia [ editar ]
.jpg)
En la literatura más antigua, la adenina a veces se llamaba vitamina B 4. [5] Debido a que es sintetizado por el cuerpo y no es esencial obtenerlo con una dieta, no cumple con la definición de vitamina y ya no forma parte del complejo de vitamina B. Sin embargo, dos vitaminas B, niacinay riboflavina , se unen a la adenina para formar los cofactores esenciales nicotinamida adenina dinucleótido (NAD) y flavina adenina dinucleótido(FAD), respectivamente. Hermann Emil Fischer fue uno de los primeros científicos en estudiar adenina.
Fue nombrado en 1885 por Albrecht Kossel , en referencia al páncreas (una glándula específica , en griego , " ν "aden") de la cual se extrajo la muestra de Kossel. [6] [7]
Los experimentos realizados en 1961 por Joan Oró han demostrado que se puede sintetizar una gran cantidad de adenina a partir de la polimerización del amoníaco con cinco moléculas de cianuro de hidrógeno (HCN) en solución acuosa; [8] si esto tiene implicaciones para el origen de la vida en la Tierra está en debate. [9]
El 8 de agosto de 2011, se publicó un informe, basado en estudios de la NASA con meteoritos encontrados en la Tierra , que sugiere que los bloques de construcción de ADN y ARN (adenina, guanina y moléculas orgánicasrelacionadas ) pueden haberse formado de manera extraterrestre en el espacio exterior . [10] [11] [12] En 2011, los físicos informaron que la adenina tiene un "rango inesperadamente variable de energías de ionización a lo largo de sus vías de reacción", lo que sugiere que "comprender los datos experimentales sobre cómo la adenina sobrevive a la exposición a la luz UVes mucho más complicado de lo que se pensaba "; estos hallazgos tienen implicaciones para las mediciones espectroscópicas de compuestos heterocíclicos , según un informe.
Un alelo ( / ə l i l / ) [1] [2] es una forma variante de un determinado gen . [3] A veces, la presencia de diferentes alelos del mismo gen puede dar lugar a diferentes rasgos fenotípicos observables , como la pigmentacióndiferente . Un ejemplo notable de este rasgo de variación de color es el descubrimiento de Gregor Mendel de que los colores de las flores blancas y moradas en las plantas de guisantes fueron el resultado de rasgos de "línea pura" que podrían usarse como control para futuros experimentos. Sin embargo, la mayoría de las variaciones genéticas dan como resultado poca o ninguna variación observable.
La mayoría de los organismos multicelulares tienen dos conjuntos de cromosomas ; Es decir, son diploides . En este caso, los cromosomas se pueden emparejar: cada par está formado por dos cromosomas del mismo tipo, conocidos como cromosomas homólogos . Si ambos alelos en un gen (o locus ) en los cromosomas homólogos son iguales, ellos y el organismo son homocigotos con respecto a ese gen (o locus). Si los alelos son diferentes, ellos y el organismo son heterocigotos con respecto a ese gen.
Etimología [ editar ]
La palabra "alelo" es una forma corta de alelomorfo ("otra forma", acuñada por los genetistas británicos William Bateson y Edith Rebecca Saunders ), [4] [5] que se usó en los primeros días de la genética para describir formas variantes De un gen detectado como fenotipos diferentes . Deriva del prefijo griego ἀλληλο-, allelo- , que significa "mutuo", "recíproco", o "entre sí", que a su vez está relacionado con el adjetivo griego ἄλλος, allos (relacionado con latín alius ), que significa "otro".
Alelos que conducen a fenotipos dominantes o recesivos [ editar ]
En muchos casos, las interacciones genotípicas entre los dos alelos en un locus pueden describirse como dominantes o recesivas , según cuál de los dos fenotipos homocigotos más heterocigotos se parece más. Cuando el heterocigoto es indistinguible de uno de los homocigotos, el alelo expresado es el que conduce al fenotipo "dominante", [6] y se dice que el otro alelo es "recesivo". El grado y el patrón de dominancia varían entre los loci. Este tipo de interacción fue descrito formalmente por primera vez por Gregor Mendel . Sin embargo, muchos rasgos desafían esta simple categorización y los fenotipos se modelan por co-dominancia y herencia poligénica .
El término alelo " tipo salvaje " se usa a veces para describir un alelo que se cree que contribuye al carácter fenotípico típico como se ve en las poblaciones de organismos "salvajes", como las moscas de la fruta ( Drosophila melanogaster ). Tal alelo de "tipo salvaje" se consideró históricamente como un fenotipo dominante (abrumador - siempre expresado), común y normal, en contraste con los alelos " mutantes " que conducen a fenotipos recesivos, raros y con frecuencia nocivos. Anteriormente se pensaba que la mayoría de los individuos eran homocigotos para el alelo de "tipo salvaje" en la mayoría de los loci genéticos, y que cualquier alelo alternativo "mutante" se encontraba en forma homocigótica en una pequeña minoría de individuos "afectados",, y más frecuentemente en forma heterocigótica en " portadores " para el alelo mutante. Ahora se aprecia que la mayoría o todos los loci de los genes son altamente polimórficos, con múltiples alelos, cuyas frecuencias varían de una población a otra, y que una gran cantidad de variación genética está oculta en forma de alelos que no producen diferencias fenotípicas obvias.
Alelos múltiples [ editar ]





Una población o especie de organismos incluye típicamente alelos múltiples en cada locus entre varios individuos. La variación alélica en un locus se puede medir como el número de alelos ( polimorfismo ) presente, o la proporción de heterocigotos en la población. Un alelo nulo es una variante genética que carece de la función normal del gen porque no se expresa o la proteína expresada está inactiva.
Por ejemplo, en el locus genético de los antígenos de carbohidratos de tipo sanguíneo ABO en humanos, [7] la genética clásica reconoce tres alelos, I A , I B e i, que determinan la compatibilidad de las transfusiones de sangre. Cualquier individuo tiene uno de los seis genotiposposibles (I A I A , I A i, I B I B , I B i, I A I B y ii) que producen uno de los cuatro fenotipos posibles : "Tipo A" (producido por Yo un yo un homocigotos y genotipos heterocigotos I A i), "Tipo B" (producido por I B I B genocigotos homocigotos e I Bi heterocigotos), "Tipo AB" producido por I A I B genotipo heterocigótico, y "Tipo O" producido por ii genotipo homocigoto. (Ahora se sabe que cada uno de los alelos A, B y O es en realidad una clase de alelos múltiples con diferentes secuencias de ADN que producen proteínas con propiedades idénticas: más de 70 alelos se conocen en el locus ABO. [8] De aquí que el individuo con sangre "Tipo A" puede ser un heterocigoto AO, un homocigoto AA o un heterocigoto AA con dos alelos "A" diferentes).
Genotipo frecuencias [ editar ]
La frecuencia de alelos en una población diploide puede usarse para predecir las frecuencias de los genotipos correspondientes (ver el principio de Hardy-Weinberg ). Para un modelo sencillo, con dos alelos;


donde p es la frecuencia de un alelo y q es la frecuencia del alelo alternativo, que necesariamente se suma a la unidad. Luego, p 2 es la fracción de la población homocigótica para el primer alelo, 2 pq es la fracción de heterocigotos y q 2 es la fracción homocigótica para el alelo alternativo. Si el primer alelo es dominante al segundo, entonces la fracción de la población que mostrará el fenotipo dominante es p 2 + 2 pq , y la fracción con el fenotipo recesivo es q 2 .
Con tres alelos:
- y


En el caso de alelos múltiples en un locus diploide, el número de genotipos posibles (G) con un número de alelos (a) viene dado por la expresión:

Dominancia alélicas en trastornos genéticos [ editar ]
Se producen varios trastornos genéticos cuando un individuo hereda dos alelos recesivos para un rasgo de un solo gen. Los trastornos genéticos recesivos incluyen albinismo , fibrosis quística , galactosemia , fenilcetonuria(PKU) y enfermedad de Tay-Sachs . Otros trastornos también se deben a alelos recesivos, pero como el locus del gen está ubicado en el cromosoma X, por lo que los hombres solo tienen una copia (es decir, son hemicigotos), son más frecuentes en hombres que en mujeres. Los ejemplos incluyen ceguera al color rojo-verde y síndrome de X frágil .
Otros trastornos, como la enfermedad de Huntington , ocurren cuando un individuo hereda solo un alelo dominante.
Epialelos [ editar ]
Mientras que los rasgos hereditarios se estudian típicamente en términos de alelos genéticos, las marcas epigenéticas , como la metilación del ADN, pueden heredarse en regiones genómicas específicas de ciertas especies, un proceso denominado herencia epigenética transgeneracional . El término epialele se usa para distinguir estas marcas hereditarias de los alelos tradicionales, que se definen por secuencia de nucleótidos . [9]Se ha descubierto una clase específica de epialele, los epialeleles metaestables , en ratones y en humanos, que se caracteriza por el establecimiento estocástico (probabilístico) del estado epigenético que puede heredarse mitóticamente.
No hay comentarios:
Publicar un comentario