| Cromatina » | |||||||||
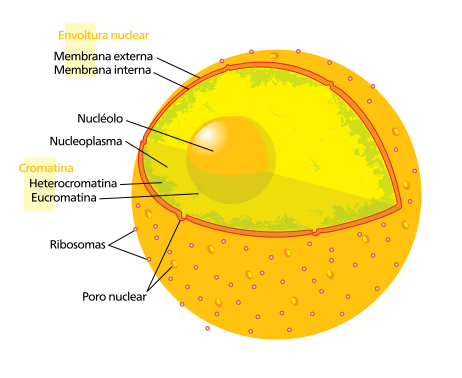
Los poros nucleares son complejos proteicos localizados en la envuelta nuclear cuya misión es permitir y controlar el tráfico de moléculas entre el nucleoplasma y el citoplasma.
Las proteínas que forman los poros nucleares se denominan nucleoporinas. El transporte mediado por los poros nucleares es específico y aprovecha el gradiente creado por las moléculas Ran-GTP y Ran-GDP. Las importinas y las exportinas son dos de las familias de moléculas que reconocen péptidos señal de entrada o salida, respectivamente, de aquellas moléculas que tienen que entrar o salir del núcleo. Son estas dos familias de proteínas las que interaccionan con las nucleoporinas durante su trasiego por los poros nucleares.
Las cisternas de la envuelta nuclear, compuestas por una membrana interna, una externa y un espacio entre ambas, dejan unos huecos entre ellas donde se encuentran los poros nucleares. Éstos son grandes complejos moleculares visibles con el microscopio electrónico y denominados en su conjunto complejo del poro. Son la puerta de comunicación entre núcleoplasma y citoplasma, y todo el transporte entre ambos compartimentos se da a través de los poros nucleares. Son un elemento clave en la función, respuesta a señales externas o diferenciación de las células. Y esto es así porque condicionan, por ejemplo, la salida del ARNm al citoplasma, o la entrada al núcleo de los factores de transcripción que determinan la expresión génica.
Los poros nucleares son muy numerosos en las células que requieren un alto tránsito de sustancias entre el núcleo y el citoplasma como, por ejemplo, en las células que se están diferenciando. Se estima que puede haber unos 11 poros por µm2 de envuelta nuclear, lo que equivale a unos 3000 a 4000 poros por núcleo.

Esquema de la estructura proteica de los poros nucleares. (Modificado de Beck et al., 2007)
Las proteínas que forman parte del complejo del poro se denominannucleoporinas. En las levaduras hay unas 30 nucleoprinas distintas en cada poro nuclear, mientras que en los metazoos pueden ser 40 o más. Se cree que un poro de una célula de mamíferos contiene unas 400 nucleoporinas, ya que muchas de ellas se encuentran repetidas. Las proteínas que forman los poros nucleares se asocian para formar 8 bloques que configuran un octágono regular y se organizan formando anillos: el anillo citoplasmático orientado hacia el citoplasma, el anillo radial situado en el hueco que deja la envuelta nuclear y es responsable de anclar el complejo del poro a las membranas de la envuelta nuclear, y el anillo nuclear que se encuentra hacia el nucleoplasma. Además, desde cada bloque, en los anillos citoplasmáticos y nuclear, se proyectan fibrillas proteicas que van hacia el citoplasma denominadas fibras citoplasmáticas, y otras al interior del núcleo que reciben el nombre de fibras nucleares. Éstas últimas se conectan a otro conjunto de proteínas que forman una estructura cerrada llamada anillo distal. Ambos, fibras nucleares y anillo distal forman la jaula nuclear. Hay que tener en cuenta que el poro en su conjunto no es una estructura transmembrana como ocurre con proteínas como los canales, aunque algunas nucleoporinas sí lo sean, sino que se podría considerar como un complejo proteico enorme anclado a las membranas de la envuelta nuclear.

Imagen tomada con un microscopio electrónico de transmisión. Se observa la envuelta nuclear con dos constricciones que se corresponden con dos poros nucleares.
Los poros nucleares contienen un pasaje acuoso interno de unos 80 a 90 nm de diámetro, pero el espacio útil para el trasiego de las moléculas que se transportan es de unos 45 a 50 nm de diámetrocuando está en reposo, y se puede expandir cuando realizan transporte activo. Normalmente, este canal permite el paso libre de pequeñas moléculas (menores de 60 kDa) pero restringe el movimiento de otras de mayor tamaño, con un potencial papel fisiológico. Aunque incluso algunas moléculas menores de 20-30 kDa tales como las histonas, los ARNt o algunos ARNm son transportadas por mediación de las nucleoporinas. Las moléculas que cruzan gracias a las nucleoporinas lo hacen por transporte pasivo facilitado. Aunque sea pasivo facilitado viajan en contra de su gradiente de concentración. Por tanto necesitan energía, la cual es aportada por el gradiente de otras moléculas denominadas Ran-GTPasas. Veamos como ocurre.

Gradientes creados por las moléculas Ran entre el citoplasma y el nucleoplasma. La energía se consume en el nucleoplasma para crear GTP y unirlo moléculas Ran para crear Ran-GTP , manteniendo la concentración de Ran-GTP elevada. En el citoplasma las Ran-GTP son rápidamente convertidas en Ran-GDP, manteniendo la concentración de estas últimas elevada. Mientras, la hidrólisis del Ran-GDP a Ran más GDP mantiene la concentración de Ran-GDP baja en el nucleoplasma. El tamaño de los iconos son indicativos de la concentración.
El transporte mediado por los poros nucleares está orquestado por las proteínas Ran y por unas familias de proteínas denominadas importinas y exportinas. Las moléculas Ran son trascendentales tanto para la importación como para la exportación de moléculas. Son las responsables de crear un gradiente que dirige el transporte, y crear este gradiente es la única parte del transporte a través de los poros nucleares que gasta energía. Las moléculas Ran pueden estar en tres estados: Ran-GTP, Ran-GDP y Ran. El paso de un estado a otro está mediado por otras enzimas. En el nucleoplasma hay una mayor concentración de Ran-GTP, mientras que en el citoplasma abunda la Ran-GTP.
Las proteínas que tienen que ser importadas al nucleoplasma poseen una secuencia de aminoácidos denominada péptido señal de entrada y las que tienen que ser exportadas un péptido señal de salida. Estas secuencias de aminoácidos serán reconocidas por las importinas o por las exportinas, respectivamente. Las proteínas de los poros nucleares no interaccionan directamente con las proteínas transportadas sino son las importinas y las exportinas. Estos complejos importina-proteína o exportina-proteína utilizan los gradientes de las moléculas Ran-GTP o Ran-GDP para cruzar los poros nucleares y llevar sus cargas al otro lado. Además de proteínas, las moléculas de ARN deben también atravesar los poros nucleares. El mecanismo que usan los distintos tipos de ARN para ser transportados difieren entre sí, pero todos están mediados por un mecanismo de asociación con proteínas. El exporte de ARNt sigue un mecanismo en el que es reconocido por una exportina denominada exportina-t, que también se une a una Ran-GTP. El mecanismo de exportación de los ARNr no se conoce muy bien. Los ARNm no utilizan, en su mayoría el sistema de transporte mediado por Ran-GTP sino oro mediado por dos proteínas que forman el complejo Tap/Nxt, el cual interacciona con las nucleoporinas y posibilita el transporte con gasto de ATP. Una pequeña cantidad de ARNm parece usar la proteínas Crm1, siendo en este caso un transporte dependiente del gradiente creado por las proteínas Ran.
Con el microscopio electrónico de transmisión se observa que la distribución de la heterecromatina periférica se interrumpe en las proximidades del los poros nuclerares. Así, se considera a los poros nucleares como lugar de permisividad para la expresión de muchos genes inducibles. Cosa lógica puesto que son la puerta de salida de los ARN mensajeros. Este efecto parece deberse a una interacción directa de las nucleoporinas con la cromatina.
Poro nuclear:
La envoltura nuclear está interrumpida por numerosos poros nucleares que constituyen los sitios de intercambio nucleo-citoplásmicos.
Los mecanismos de intercambio nucleo-citoplásmicos de realizan en dos sentidos.
Las moléculas pequeñas (nucleótidos, proteínas...) de peso molecular inferior a 40 kDa y los iones atraviezan el poro por difusión.
El transporte de moléculas grandes se produce con la participación de proteínas asociadas y proteínas del complejo del poro.
¿Cuáles moléculas son importadas del citosol al nucleoplasma?
Las principales son: -Enzimas para la transcripción y replicación del ADN.
-Proteínas que forman parte de las sub-unidades ribosomales.
-Proteínas que regulan la transcripción de genes.
-Complejos formados por Hormonas esteroideas y sus receptores.
-Moléculas de la familia Hsp (Heat Shock Protein) o "chaperonas".
-Proteínas virales (en caso de infección viral).
¿Cuáles moléculas son exportadas del núcleo al citoplasma? Las principales son: -ARN r (sub-unidades ribosomales)
-ARN t
-ARN m
-
| |||||||||




http://www.medic.ula.ve/histologia/anexos/celulavirtual/nucleo/poro.htm
Los "poros nucleares" son grandes complejos de proteínas que atraviesan la envoltura nuclear, la cual es una doble membrana que rodea al núcleo celular, presente en la mayoría de los eucariontes. Hay cerca de 3000 a 4000 Complejos de Poro Nuclear (ingl:NPC) en la envoltura nuclear de la célula de un vertebrado, pero su número varía dependiendo del número de transcripciones de la célula. Las proteínas que forman los complejos de poro nucleares son conocidas como nucleoporinas. Cerca de la mitad de las nucleoporinas contienen comúnmente una estructura terciaria alfa solenoideo beta hélice, o en algunos casos ambas como dominios proteicos separados. La otra mitad muestra características estructurales típicas de proteínas "nativamente no dobladas", por ejemplo son proteínas altamente flexibles que carecen de estructura secundaria ordenada.1 Estas proteínas desordenadas son las nucleoporinas FG, llamadas así por su secuencia aminoacídica que contiene varias repeticiones del péptido fenilalanina-glicina2 .
Los poros nucleares permiten el transporte de moléculas solubles en agua a través de la envoltura nuclear. Este transporte incluye el movimiento de ARN y ribosomas desde el núcleo al citoplasma, y movimiento de proteínas (tales como ADN polimerasas y laminas), carbohidratos, moléculas de señal y lípidos hacia el núcleo. Es notable que el complejo de poro nuclear (CPN o NPC en inglés) puede conducir activamente 1000 translocaciones por complejo por segundo. Aunque las moléculas pequeñas pasan por difusión simple a través de los poros, las moléculas de mayor tamaño pueden ser reconocidas mediante secuencias de señal específicas y luego difundidas con la ayuda de las nucleoporinas hacia o desde el núcleo. Esto es conocido como el ciclo RAN. Cada una de las ocho subunidades proteicas que rodean el poro verdadero (el anillo externo) proyecta una proteína con forma de radio hacía el canal del poro. El centro del poro muchas veces parece que tuviera una estructura parecida a un tapón. Aún no se sabe sí esto corresponde a un tapón verdadero o es simplemente carga atrapada durante el tránsito.
No hay comentarios:
Publicar un comentario