Las células se adhieren a estructuras próximas principalmente mediante proteínas transmembrana localizadas en la membrana plasmática.
La adhesión de las células a la matriz extracelular está mediada sobre todo por integrinas.
La adhesión directa entre células está mediada por cadherinas, inmunoglobulinas, selectinas e integrinas.
La adhesión de la célula tiene otras funciones, además de la sujetar, y actúa como un mecanismo de transducción que le permite a la célula conocer su entorno extracelular. Cambios en la adhesión afectan a la fisiología celular y puede desencadenar la expresión génica.
Las células de los organismos pluricelulares se organizan en tejidos y órganos. Esta disposición depende en gran medida de su capacidad para adherirse bien a la matriz extracelular o a otras células. La adhesión no sólo sirve para anclar y situar a las células para formar andamiajes tridimensionales, sino también para comunicarse entre sí. Es decir, el grado de adhesión y a quién se adhieren las células es un tipo de información útil para la célula. La adhesión se realiza por medio de las denominadas proteínas de adhesión, las cuales se encuentran ancladas a la membrana plasmática. Hay dos tipos, aquellas que anclan la célula a la matriz extracelular y aquellas que stablecen uniones directas entre dos células contiguas.
Adhesión de la célula de la matriz extracelular.
 Más que adhesión
Las integrinas son las moléculas más importantes en la adhesión de la célula a la matriz extracelular. Son una gran familia de proteínas presentes en prácticamente todos los animales. Son proteínas transmembrana formadas por dos subunidades (alfa y beta). En mamíferos hay 18 unidades alfa y 3 unidades beta que por combinación pueden formar hasta 24 integrinas diferentes, las cuales se expresan según el tejido o estado fisiológico de la célula. Cada una tiene 3 dominios moleculares: un dominio intracelular que contacta con los filamentos de actina del citoesqueleto, otro extracelular globular que es capaz de unirse al colágeno, fibronectinas y lamininas, y un dominio transmembrana entre las cadenas de ácidos grasos de la membrana. Su capacidad de unirse a moléculas de la matriz extracelular y al citoesqueleto permite una continuidad estructural mecánica entre el interior y exterior de la célula. Pero además permite modificar el comportamiento celular en función de las moléculas presentes en la matriz extracelular. Esto es posible porque el estado de adhesión de la integrina se transmite a su dominio citosólico, el cual interactúa con proteínas que son capaces de viajar al interior del citoplasma para afectar a rutas moleculares o viajar al interior del núcleo para alterar la expresión génica. También la célula puede modificar su capacidad de adhesión, y por tanto su movilidad, cambiando el juego de proteínas de adhesión en su superficie ☆.

Integrinas en diferentes estado de activación (Modificado de Luo et al., 2007)
Las integrinas suelen aparecer asociadas en la membrana plasmática formando las denominadas adhesiones focales y también formar agregados mayores como son los hemidesmosomas. La fuerza de unión de una célula a la matriz extracelular depende pues de la cantidad, tipo y estado de las integrinas que presenta en su membrana plasmática.

Principales proteínas transmembrana que realizan contactos célula-célula (Modificado de Hynes, 1999)
Adhesión entre células.
Estas moléculas se encargan de adherir directamente unas células a otras. Hay cuatro tipos: cadherinas, inmunoglobulinas, selectinas y algunos tipos de integrinas. Las cadherinas se encuentran en la superficie de la mayoría de las células animales y forman uniones homotípicas, es decir, reconocen a otras cadherinas en la célula adyacente. Son una gran superfamilia de proteínas cuyos miembros suelen aparecer característicamente en ciertos tejidos. Así, la N-cadherina se expresa en el tejido nervioso, la E-cadherina en el tejido epitelial, etcétera. Es por ello que juegan un papel importante en la segregación de poblaciones celulares de los distintos tejidos. Son especialmente importantes durante el desarrollo embrionario. Las cadherinas son parte estructural de los desmosomas (macula adherens). Las moléculas de adhesión del tipo inmunoglobulina, también llamadas CAM (cell adhesion molecule) forman uniones homofílicas con inmunoglobulinas presentes en la célula adyacente, aunque también pueden realizar uniones heterofílicas con otro tipo de moléculas. Es también una gran familia de proteínas con distribución específica de sus miembros en los distintos tejidos. Por ejemplo, N-CAM aparece en el sistema nervioso. Sus uniones no son tan fuertes como las de las cadherinas y parece que actúan ajustando de forma más fina la asociación entre células de un mismo tejido. Las selectinas son también proteínas de adhesión entre células, pero forman uniones heterofílicas, es decir, se unen a moléculas de diferente tipo de la otra célula, en concreto a glúcidos presentes en la célula vecina. Esto es gracias a que poseen un dominio que tiene apetencia por determinados azúcares (ácido siálico y fucosa). Son importantes en la unión de los glóbulos blancos a las paredes del endotelio cuando abandonan el torrente sanguíneo para adentrarse en los tejidos. Las integrinas, que antes vimos como moléculas que median la adhesión de las células a la matriz extracelular, también pueden mediar adhesiones célula-célula. En concreto, algunas integrinas pueden formar uniones con algunas moléculas transmembrana del tipo de las inmunoglobulinas.
Membrana celular.
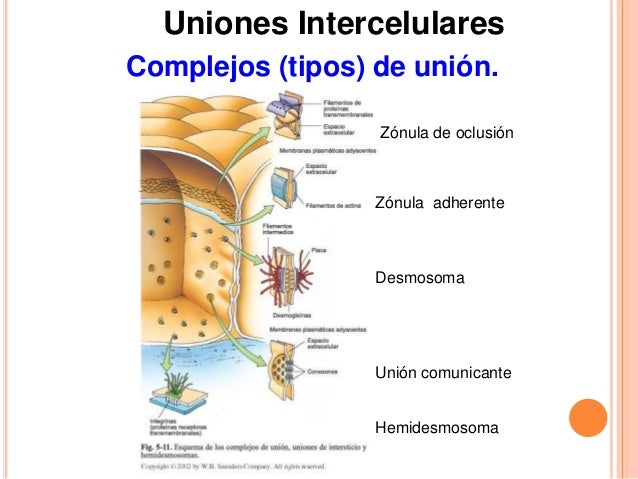
COMPLEJOS de UNIÓN
Existen uniones entre células que forman estructuras macromoleculares denominados complejos de unión.
Uniones estrechas. Forman asociaciones muy fuertes entre células, obliterando el espacio intercelular a modo de cinturón en todo el perímetro celular.
Uniones adherentes. Aparecen en las células epiteliales a modo de cinturón, en todo el perímetro celular, importantes durante la morfogénesis.
Desmosomas. Forman uniones puntuales a modo de remaches. Aparecen en numerosos tejidos.
Hemidesmosomas. Forman uniones puntuales entre la membrana basal de las células epiteliales y la lámina basal.
Uniones focales. Concentraciones de moléculas en zonas de la membrana plasmática que forman uniones con la matriz extracelular.
Como hemos visto en el apartado anterior las células se anclan a la matriz extracelular y a otras células mediante unas proteínas especializadas. Las integrinas, cadherinas, selectinas e inmunoglobulinas son las más importantes. A veces se producen uniones tan especializadas y desarrolladas que forman estructuras macromoleculares denominadas complejos de unión y uniones focales, las cuales son fundamentales para mantener la cohesión de muchos tejidos, principalmente los epitelios, el tejido muscular y el nervioso.
Los complejos de unión se clasifican según su forma, las moléculas de adhesión que los componen, los elementos a los que se unen y sus interacciones con el citoesqueleto. La primera vez que se observaron fue con el microscopio electrónico y se clasificaron morfológicamente, pero fueron las técnicas de biología molecular las que permitieron desentrañar sus estructuras moleculares.

Esquema de las uniones estrechas de las células epiteliales del digestivo. La estructura molecular parece ser similar en los distintos tipos de epitelios. (Modificado de Niessen 2007).
Las uniones estrechas o zonula occludens se encuentran en diferentes tipos celulares, como en las partes apicales de los epitelios, en los endotelios del sistema nervioso, o en el tejido muscular cardiaco. Establecen uniones tan fuertes y estrechas entre las células contiguas que prácticamente no dejan espacio intercelular entre sus membranas plasmáticas, limitando la difusión de sustancia solubles extracelulares.
En el caso de las células epiteliales forman una especie de cinturón que rodea todo el perímetro celular. Además de mantener cohesionadas fuertemente a las células realizan otras funciones. En los epitelios, por ejemplo en el epitelio digestivo, impiden la difusión intercelularevitando que las sustancias del interior del tubo digestivo penetren en el organismo por los espacios intercelulares. Esto obliga a las sustancias a ser captadas selectivamente por parte de las células epiteliales, donde son transformadas y liberadas al torrente sanguíneo. Pero, además, las uniones estrechas permiten la polaridad de las células epiteliales puesto que impiden la difusión lateral de moléculas insertas en sus membranas celulares. Es decir, actúan como una barrera física a la difusión lateral de las moléculas de la membrana plasmática. Con ello se consigue una zona o dominio apical con un juego de moléculas distinto al que hay en el domino latero-basal de la célula epitelial. Esta separación es importante para establecer un camino de captación y liberación de sustancias desde el exterior hacia el interior.
En los capilares del sistema nervioso central las células endoteliales están unidas por uniones estrechas que contribuyen a establecer la barrera hematoencefálica, la cual es un filtro importante para las moléculas que tienen intercambiarse entre la sangre y las neuronas y glía
Las uniones estrechas están formadas por la ocludina y por una familia de moléculas denominadas claudinas, que son las proteínas transmembrana encargadas de establecer los contactos célula-célula. Las claudinas parecen ser las más importantes en el establecimiento de la unión y en estas uniones forman unos poros que dejan pasar ciertos iones por el espacio extracelular, no más de 1 nanometro de diámetro. Hay 20 tipos de claudinas, cada una de las cuales forma uno poro extracelular distinto y así los epitelios pueden modificar la selectividad de su permeabilidad intercelular según el tipo de claudina que expresen. El dominio intracelular de estas moléculas interactúa con otras moléculas denominadas ZO, las cuales forman un entramado que interacciona con los filamentos de actina del citoesqueleto y con otras proteínas citosólicas que desencadenan cascadas de señalización. Una observación curiosa es que las uniones estrechas parecen depender de la presencia de uniones adherentes. Así, cuando se impide la formación de uniones adherentes en una célula, ésta no es capaz de establecer uniones estrechas.
Las uniones adherentes o zonula adherens son complejos de unión que se forman en las células epiteliales y que se sitúan próximas y basales a las uniones estrechas. Su misión es unir células vecinas. Son los primeros complejos de unión que se forman durante el desarrollo de los epitelios, aparecen antes que las uniones estrechas, por lo que parecen actuar en procesos morfogenéticos durante el desarrollo embrionario. Al igual que las uniones estrechas forman una estructura a modo de cinturón en todo el perímetro celular. Las E-cadherinas son las moléculas encargadas de realizar las conexiones célula-célula con su dominio extracelular, mientras que el intracelular contacta con los filamentos de actina. En el entramado molecular que se asocia con el dominio interno de las cadherinas se encuentra la β-catenina, la cual puede desencadenar cambios en la expresión génica cuando se desplaza hasta el núcleo.

Organización y composición de los desmosomas (modificado de Huber 2003)
Los desmosomas o macula adherens, al contrario que los dos complejos de unión anteriores, establecen conexiones puntuales en forma de disco entre células vecinas, como si fuesen remaches. Son muy abundantesentre las células epiteliales y entre las musculares, pero también en otros tejidos como el nervioso. Las uniones entre células están mediadas por moléculas del tipo cadherinasdenominadas desmogleínas y desmocolinas. El dominio intracelular de estas cadherinas contacta con los filamentos intermedios como las queratinas, gracias a proteínas intermediarias.
Los hemidesmosomas y las uniones focales establecen uniones fuertes entre las células y la matriz extracelular. En ambos casos las uniones se establecen por integrinas. Los hemidesmosomas unen las células epiteliales a la lámina basal gracias al dominio extracelular de la integrina, mientras que el dominio intracelular contacta con los filamentos intermedioscitosólicos. Aunque los hemidesmosomas parecen desmosomas sin una de sus mitades, molecularmente son diferentes. Las uniones focales unen a las células con diversos tipos de matrices extracelulares gracias a otro tipo de integrinas que en su dominio intracelular contacta con los filamentos de actina.

Imágenes de microscopía electrónica de transmisión de la epidermis mostrando desmosomas y hemidesmosomas.

Esquema un hemidesmosma localizado en la base de un epitelio de mamífero. (Modificado de Hahn 2001)
Algunos autores suelen colocar en este apartado de estructuras cohesivas macromoleculares a las uniones en hendidura. Estas son uniones entre células establecidas por unas moléculas denominadas conexinas. Sin embargo, las uniones en hendidura no tienen como principal misión cohesionar tejidos sino permitir la comunicación directa entre citoplasmas de células vecinas, gracias a los canales que crean las conexinas. Por tanto, veremos estas estructuras cuando hablemos de la comunicación celular.
|
|



No hay comentarios:
Publicar un comentario