LA MATRIZ EXTRACELULAR
-Corresponde a los espacios existentes entre las células (individuos pluricelulares) ocupados por una gran gama de macromoléculas.
-Es sintetizada por las propias células durante la embriogénesis y persiste durante toda la vida del organismo.
-Funciones más destacables:
a)dar soporte y rigidez a las células y tejidos.
b)mantener las células unidas y comunicadas entre sí.
c)influir en el metabolismo celular.
d)actuar sobre la organización del citoesqueleto, a través de la membrana plasmática.
-La composición macromolecular de la matriz es muy variada: en los vertebrados se puede endurecer para originar, por ejemplo, los huesos o los dientes; en los vegetales forma la pared celular.
Las principales macromoléculas que constituyen la matriz son éstas:
1)Polisacáridos que, unidos a proteínas, forman proteoglucanos. Forman geles hidratados, entre los cuales se halla el ác. hialurónico, cuya principal función es facilitar la migración de las células.
2)Proteínas estructurales, como el colágeno y la elastina.
LA PARED CELULAR
-Es una cubierta extracelular que rodea a las células vegetales (con la excepción de los gametos de las plantas superiores).
-Características:
a)es un exoesqueleto que protege, da rigidez y soporte a la planta.
b)tiene un espesor variable, dependiendo del tipo y función de los tejidos.
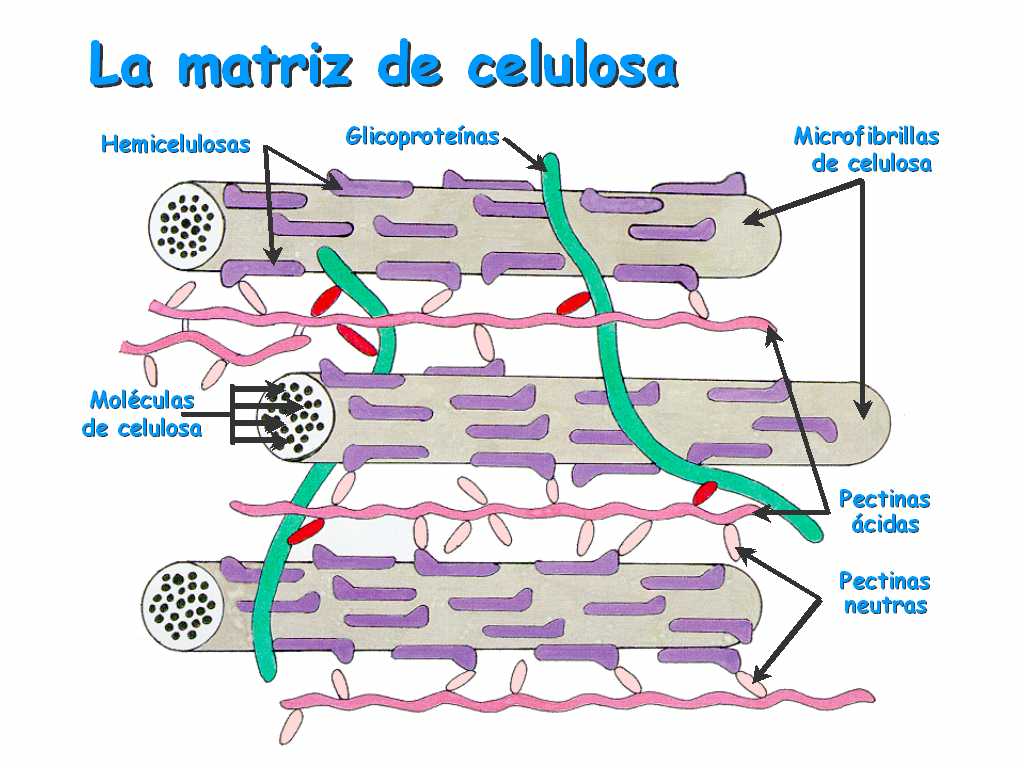
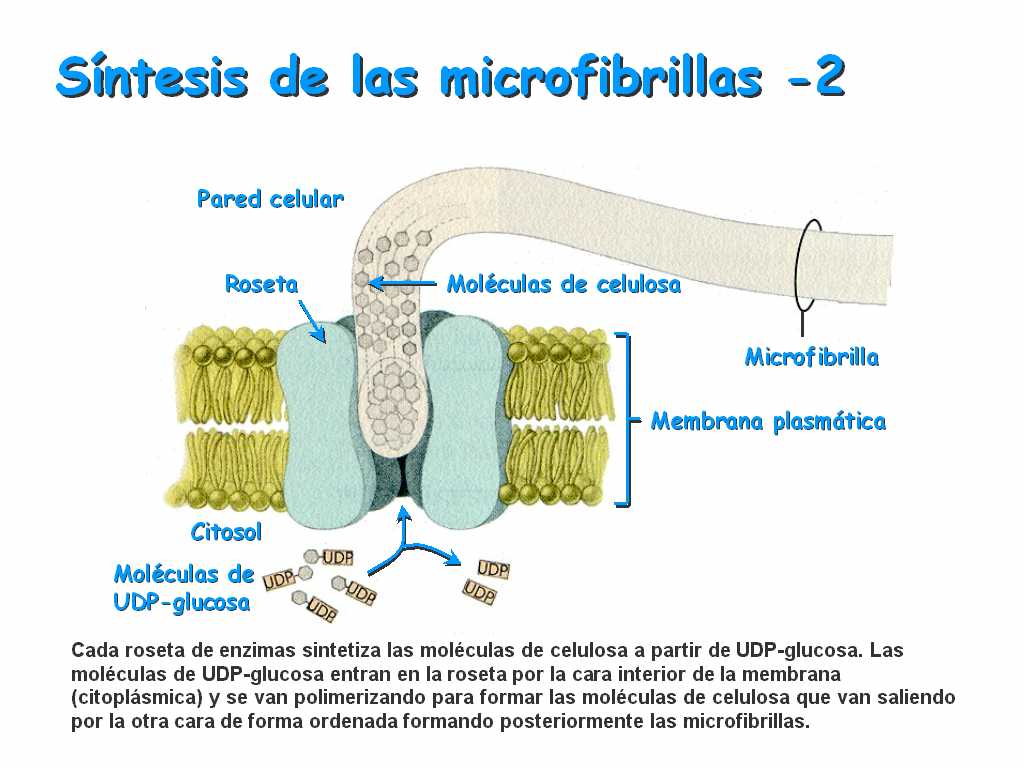
c)está compuesta por fibras de celulosa embebidas en polisacáridos y proteínas.
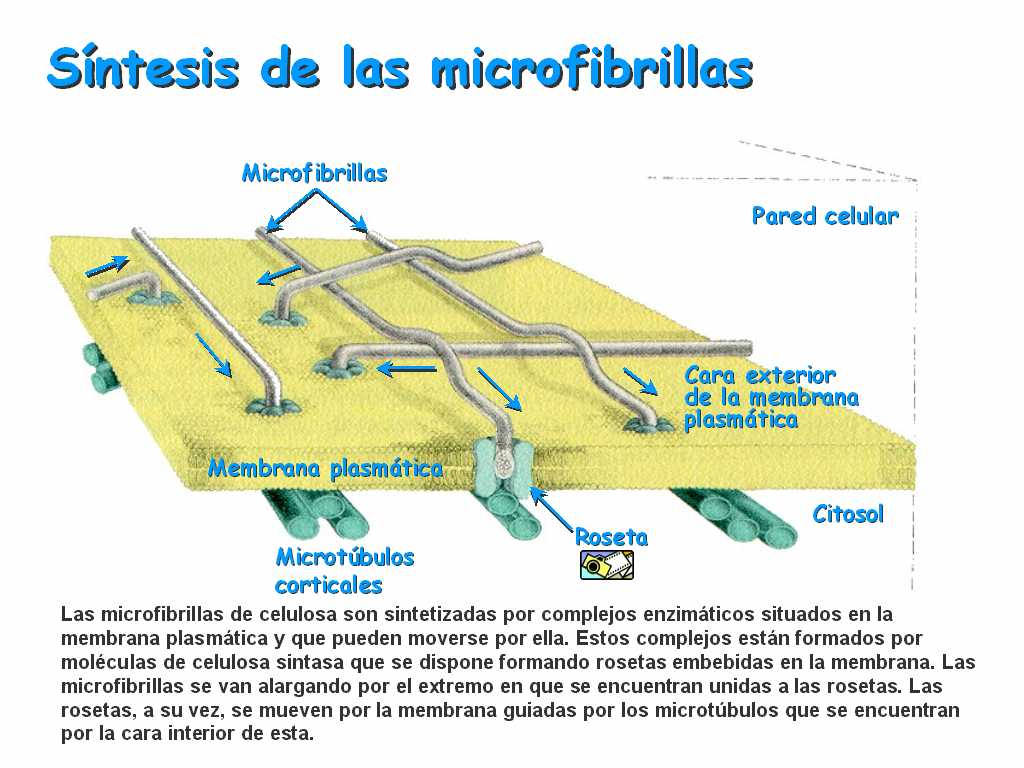
d)la celulosa se forma en el aparato de Golgi, aunque también la origina un complejo enzimático de la membrana (la celulosintetasa).

Organización de la pared celular
-Varía según el tipo de tejido y su grado de diferenciación.
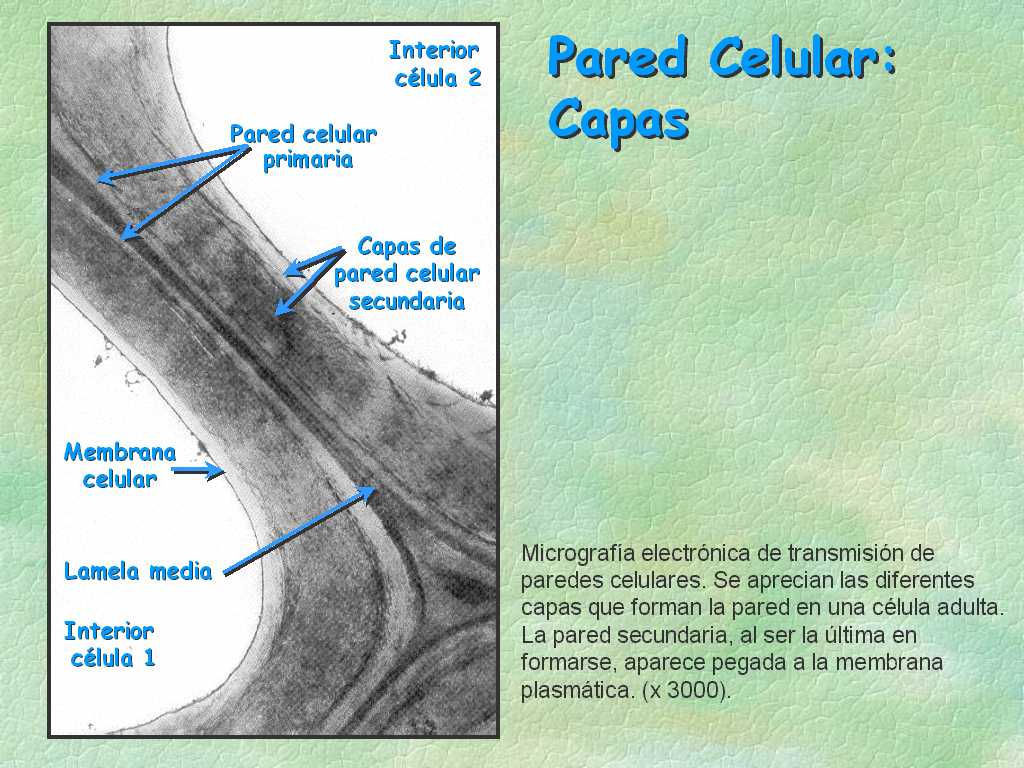
En las células recién formadas, que acaban se sufrir una mitosis, se deposita sobre la membrana plasmática una lámina, llamada lámina media, formada sobre todo por pectina y lignina.
-Cuando la célula se va diferenciando y creciendo, se van depositando de manera estratificada otros materiales sobre la lámina media: celulosa, otros polisacáridos y proteínas, así como gran cantidad de agua (80-90%). Estos materiales conforman la pared primaria.
-Cuando la célula alcanza su máximo desarrollo, la pared aumenta de grosor y se convierte en la pared definitiva. Puede ocurrir que se depositen nuevas capas (hacia dentro), que darán mayor resisitencia y grosor a las células, constituyendo la pared secundaria (esto sucede, por ejemplo, en el esclerénquima y en el xilema).
-A pesar de estas capas, la célula vegetal no se aísla de sus vecinas, ya que mantiene puentes de comunicación intercelular originados durante la telofase: son los plasmodesmos. También se producen punteaduras que corresponden a adelgazamientos de la pared celular. Las punteaduras de una célula se corresponden con las de las adyacentes.
http://perso.wanadoo.es/sancayetano2000/biologia/apu/tema2_5.htm
La matriz extracelular es un invento de los organismos pluricelulares. Es esencial para mantener a las células unidas puesto que permite la adhesión de las células para formar tejidos. Pero con el tiempo ha adquirido muchas más funciones: aporta propiedades mecánicas a los tejidos (tanto en animales como en vegetales), mantiene la forma celular, permite la comunicación intercelular, forma sendas por las que se mueven las células, modula la diferenciación y la fisiología celular, secuestra factores de crecimiento, etcétera. La cantidad, la composición y la disposición de la matriz extracelular depende del tipo de tejido considerado. Hay algunos como el epitelial y el nervioso que tienen muy poca matriz extracelular, mientras que en otros, como el tejido conectivo propiamente dicho, el cartílago o el hueso, es el elemento más importante en volumen. La composición molecular de la matriz extracelular es típica de cada tejido y sus componentes son renovados continuamente por las células que la producen. Esto supone que la matriz extracelular está en constante renovación.
Las células interaccionan con la matriz celular mediante proteínas transmembrana, principalmente las integrinas, las cuales se adhieren o reconocen a moléculas de la matriz extracelular.
En los tejidos vegetales la pared celular se puede considerar, aunque no siempre hay acuerdo, como una matriz extracelular especializada con unas características muy diferentes a la de los tejidos animales. Su papel es crucial para dar rigidez a las células y por extensión a la planta, es una barrera a la permeabilidad y protege frente a las agresiones de patógenos o mecánicas, entre otras funciones.

En esta imagen se presentan ejemplos de distintos tipos de matrices extracelulares teñidas con diferentes colorantes. Los asteriscos señalan la matriz extracelular. A) Cartílago hialino, B) Matriz ósea compacta. C) Conectivo denso regular (tendón). D) Conectivo gelatinoso del cordón umbilical. E) Paredes celulares del sistema vascular de un tallo de una planta. F) Células epiteliales. Obsérvese que prácticamente no hay sustancia intercelular. G) Imagen de microscopía electrónica del tejido nervioso donde prácticamente no existe matriz extracelular.

Esquema de las principales moléculas que aparecen en la matriz extracelular de un tejido conectivo.
Las principales macromoléculas que componen la matriz extracelular son: proteínas estructurales como el colágeno y la elastina, glicosaminoglucanos, proteoglicanos y glicoproteínas. Todas ellas se encuentran en un medio acuoso junto con otras moléculas de menor tamaño, además de iones. Es la cantidad, la proporción y el tipo de cada una de estas macromoléculas lo que distingue a unas matrices extracelulares de otras.

Imagen de microscopía electrónica de barrido de la matriz extracelular de la submucosa del digestivo Las cintas largas son fibras de colágeno.
La matriz extracelular está formada principalmente por proteínas, glicosaminoglicanos, proteoglicanos y glicoproteínas, organizados en entramados diversos que constituyen las diferentes matrices extracelulares de los distintos tejidos. Las proteínas más abundantes son el colágeno y la elastina.
Colágeno
Se denomina colágeno a una familia de proteínas muy abundante en los animales, pudiendo representar del 25 al 30 % de todas las proteínas corporales. Tradicionalmente se ha usado el colágeno para fabricar pegamentos y colas, de ahí su nombre. En los vertebrados hay más de 40 genes que sintetizan unas cadenas de aminoácidos denominadas cadenas alfa, las cuales se asocian de tres en tres para formar hasta 28 tipos de moléculas de colágeno diferentes. Su principal misión es crear un armazón que hace de sostén a los tejidos y que resiste las fuerzas de tensión mecánica. Actúa como las barras de acero que refuerzan el hormigón en los edificios. La organización de las moléculas de colágeno en estructuras macromoleculares tridimensionales es variada, pudiendo formar haces, matrices, etcétera. Las células se "agarran" a las moléculas de colágeno mediante diversas proteínas de adhesión como las integrinas, inmunoglobulinas, anexinas, etcétera.
Las moléculas de colágeno se caracterizan por:
a) Una composición poco frecuente de aminoácidos. En las moléculas de colágeno abunda el aminoácido glicina, que es muy común, y otros menos comunes como la prolina e hidroxiprolina. La glicina se repite cada 3 aminoácidos (...-Gly - x - y - Gly - x - y -...), donde x e y suelen ser prolina e hidroxiprolina, respectivamente. Esta secuencia repetida de glicina es la que permite la disposición en hélice levógira de las cadenas α, debido al pequeño tamaño de este aminoácido.

Fibras de colágeno en la matriz extracelular del tubo digestivo. Microscopía electrónica de barrido.

Fibras de colágeno de la dermis.
Tinción: tricrómico de Masson.
Tinción: tricrómico de Masson.

Fibras de colágeno en la matriz extracelular del tubo digestivo. Microscopía electrónica de trasnmisión.
b) Pueden organizarse formando fibras, mallas o especializarse en formar uniones entre moléculas. Todo ello depende de la composición química de sus subunidades α y de los tipos de subunidades que lo formen (ver tabla).
Forman fibras . Son las más abundantes de todas las formas de colágeno y están formadas por repeticiones de moléculas de colágeno, tres cadenas α arrolladas en forma de triple hélice dextrógira que forman las unidades repetidas. De los colágenos que forman fibras el más frecuente es el tipo I, que abunda en huesos, cartílago y piel, y que representa el 90 % de todo el colágeno del organismo. Otros tipos abundantes son el II, presente en el cartílago hialino, y el III, que abunda en la piel y en los vasos sanguíneos.
Forman mallas . Estos tipos de colágeno suelen organizarse en entramados moleculares que forman láminas. Se encuentran rodeando los órganos o formando la base de los epitelios. Entre éstos se encuentra el colágenotipo IV que abunda en la lámina basal, localizada entre el epitelio y el tejido conectivo.
Establecen conexiones . Forman puentes de unión entre moléculas de la matriz extracelular y el colágeno fibrilar o el colágeno que forma mallas. Por ejemplo, el colágeno tipo IX forma uniones entre los glicosaminoglicanos y las fibras de colágeno tipo II.

Esquema de la síntesis de las fibras de colágeno.
También existen moléculas de colágeno que poseen secuencias de aminoácidos hidrofóbicos y que se encuentran como moléculas transmembrana. Es el caso del colágeno tipo XIII y el tipo XVII. El colágeno tipo XVII forma parte de la estructura de los hemidesmosomas.
Independientemente del tipo, la síntesis de la moléculas de colágeno ocurre en forma de precursor. El colágeno se sintetiza en el interior celular en forma de procolágeno. En primer lugar se sintetizan las cadenas α inmaduras en el retículo, donde son modificadas: hidroxiladas las porolinas y lisinas, glucosidadas (O-glicosidación), y plegadas por chaperonas. Aquí se asocian las cadenas α de 3 en 3 mediante puentes de hidrógeno, para formar las moléculas de procolágeno. Éstas son reconocidas por receptores transmembrana y empaquetado en vesículas recubiertas por COPII. Estas vesículas han de ser especiales puesto que las moléculas de procolágeno son como varillas rígidas de unos 400 nm (las vesículas típicas COPII miden entre 60 y 90 nm). El procolágeno pasa por el aparato de Golgi, no se sabe muy bien cómo, desde donde es exocitado al exterior celular. Es destacable que algunas células pueden seleccionar el dominio celular donde se liberará un determinado tipo de colágeno. Independientemente de esto, durante, o tras la liberación, sufre una acción enzimática que elimina una secuencias terminales de cada cadena α, transformando el procolágeno en colágeno. Estas secuencias terminales impiden que el procolágeno se ensamble espontáneamente en el interior celular. Las moléculas de colágeno, sin cadenas terminales, se ensamblan automáticamente para formar las fibrillas de colágeno, que a su vez se unen para formar las fibras de colágeno (ver figura).

Imagen obtenida con un microscopio electrónico de transmisión a partir de tejido conectivo de un invertebrado marino, la oreja de mar. Con los asteriscos negros se indica el colágeno ya ensamblado en el exterior celular, mientras que con los asteriscos blancos las grandes vesículas intracelulares llenas de moléculas de procolágeno. La flecha blanca indica un posible punto de liberación de las moléculas de procolágeno al espacio extracelular.
El colágeno se sintetiza principalmente por fibroblastos, miofibroblastos, osteoblastos y condrocitos. Algunas moléculas de colágeno son también sintetizadas por otros tipos celulares tales como las epiteliales.
Elastina

Fibras de elastina del tejido conectivo
Es una proteína abundante en muchas matrices extracelulares y aparece como un componente de las denominadas fibras elásticas, las cuales son agregados insolubles de proteínas. Al contrario que las fibras de colágeno, las fibras elásticas tienen la capacidad de estirarseen respuesta a las tensiones mecánicas y de contraerse para recuperar su longitud inicial en reposo. La elasticidad de nuestros tejidos depende de las fibras elásticas. Se encuentran sobre todo en la dermis, en las paredes de las arterias, en el cartílago elástico y en el tejido conectivo de los pulmones. Además de la elastina, que representa el 90 %, las fibras elásticas están formadas por las denominadas microfibrillas de fibrilina y por otras glicoproteínas y proteoglicanos en menor proporción. Otras funciones de las fibras elásticas son aportar sostén a los tejidos o regular la actividad de los factores de crecimiento TGF-β mediado por la fibrilina.

Esquema de una porción de una fibra de elastina. Las moléculas de elastina están unidas entre sí mediante enlaces entre las regiones ricas en el aminoácido lisina (Modificado de Kielty 2007).
La elastina posee una larga cadena de aminoácidos en la que hay numerosas secuencias con aminoácidos hidrófobos, separadas por otras secuencias que contienen parejas de glicinas y otros aminoácidos pequeños como la lisina. Esta composición de aminoácidos es la que confiere las propiedades elásticas, puesto que los aminoácidos hidrófobos permiten la disposición en estructuras arrolladas y la lisina la formación de α-hélices. Los aminoácidos no hidrófobos son los puntos donde se enlanzan dos moléculas de elastina próximas. La elastina parece ser una invención de los vertebrados, puesto que no se ha encontrado en invertebrados.
http://mmegias.webs.uvigo.es/5-celulas/2-componentes_proteinas.php
LA PARED CELULAR
Pared celular
Es un componente típico de las células eucarióticas vegetales y fúngicas. Entre las Embriófitas, las únicas células que no la tienen son los gametos masculinos y a veces los gametos femeninos. En las células vivas las paredes tienen un papel importante en actividades como absorción, transpiración, traslocación, secreción y reacciones de reconocimiento, como en los casos de germinación de tubos polínicos y defensa contra bacterias u otros patógenos. Son persistentes y se preservan bien, por lo cual se pueden estudiar fácilmente en plantas secas y también en los fósiles.
Inclusive en células muertas son funcionales las paredes celulares: en los árboles, la mayor parte de la madera y la corteza está formada sólo de paredes celulares, ya que el protoplasto muere y degeneración
|
En la corteza las paredes celulares contienen materiales que protegen las células subyacentes de la desecación. En la madera las paredes celulares son gruesas y rígidas y sirven como soporte mecánico de los órganos vegetales. 
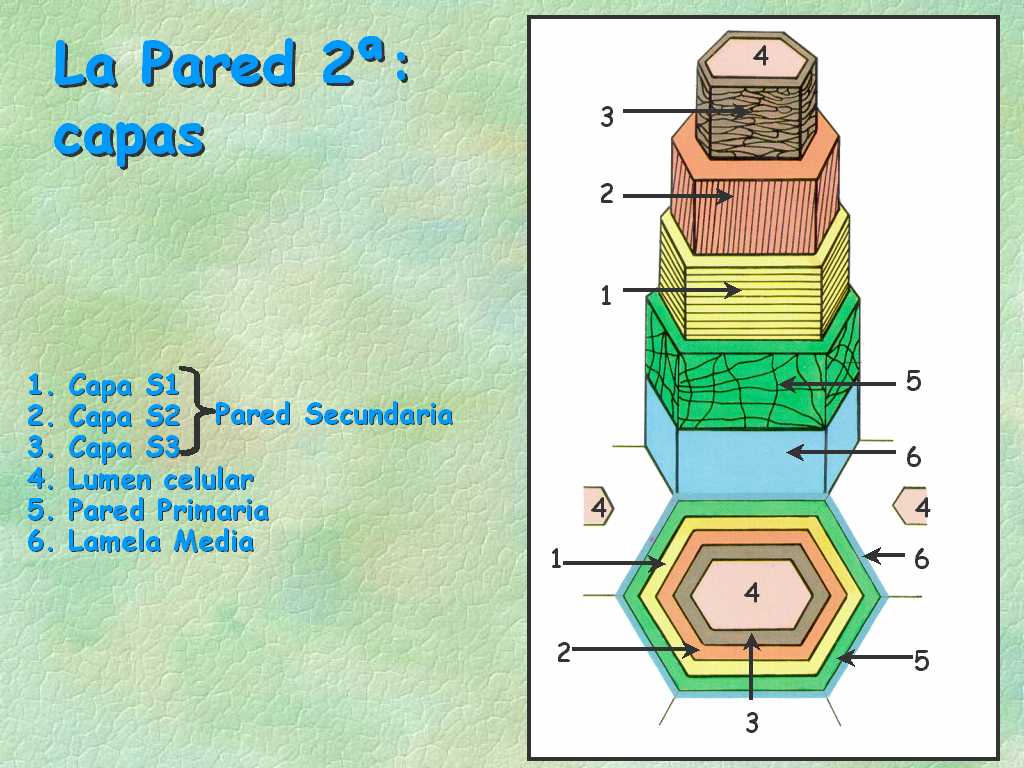
CAPAS DE LA PARED CELULAR
La pared celular tiene tres partes fundamentales: 1) la sustancia intercelular o lámina media, 2) la pared primaria y 3) la pared secundaria (Fig. 7.6). La pared es secretada por la célula viva, de manera que la capa más vieja está hacia afuera, y la capa más joven hacia adentro junto al protoplasma, demarcando el lumen o cavidad celular.
Se inicia como "placa celular", en el momento de la división celular. Es amorfay ópticamente inactiva. Se compone principalmente de compuestos pécticos (ácido péctico: unión de moléculas de ácido galacturónico; pectatos: ácido péctico + iones metálicos, como Ca o Mg). Se descompone con facilidad, y cuando ésto sucede el tejido se separa en células individuales. Ejemplos: cuando las manzanas se vuelven "harinosas" y en el proceso de "maceración".
|
Fig. 7.6. Porción de pared celular entre dos células
| ||
 |
En tejidos leñosos generalmente la laminilla media está lignificada. En los tejidos adultos la laminilla media es difícil de identificar porque se vuelve extremadamente tenue. En consecuencia, las paredes primarias de dos células contiguas y la laminilla media que se halla entre ambas con microscopio óptico se observan como una unidad que se denomina lámina media compuesta (Fig. 7.6).
Pared primaria
Se forma inmediatamente después de la división celular, antes de que la célula complete su crecimiento. Está asociada a protoplastos vivos, por lo tanto los cambios que experimenta son reversibles. Usualmente es delgada, (Fig. 7.7) pero puede alcanzar considerable grosor. Cuando las paredes son gruesas pueden mostrar una clara laminación debida a las variaciones en la composición de los sucesivos incrementos. |
Fig. 7.7. Célula vegetal con pared
primaria y granos de almidón |
Fig. 7.8, a. Fibras en transcorte
|
 |  |
Fig. 7.8, b. Fibras en transcorte de Turnera (foto MEB)
| |
 |
Pared secundaria
Sigue a la pared primaria en orden de aparición. Es fuertemente refringente al microscopio debido a la alta proporción de celulosa. La pared secundaria de traqueidas y fibras (Fig. 7.8) generalmente consta de tres capas con características físicas y químicas diferentes, que se denominan de afuera hacia adentro S1 (capa externa), S2 (capa medial o central) y S3 (capa interna). Algunos consideran que la última capa puede ser considerada como una pared terciaria, que presenta internamente una capa verrucosa, los restos de protoplasto (Fig. 7.6). En algunas células el depósito de pared no es uniforme, sino que los engrosamientos ocurren en zonas determinadas. En la pared primaria, por ejemplo en células del colénquima y en los pedúnculos de cistolitos. En la pared secundaria constituyen prominencias situadas por dentro o fuera de la célula: anillos de los elementos traqueales, apéndices y escultura de las esporas y granos de polen.
|









No hay comentarios:
Publicar un comentario