Auxesis (de la palabra griega que significa aumento; crecer) se refiere al crecimiento de un aumento en el tamaño de la célula en lugar de un aumento en el número de células. El crecimiento auxiliar se produce en ciertos tejidos, como el músculo , de los animales superiores, así como en algunos organismos, como nematodos, tunicados y rotíferos . [1] [2]
En la fisiología de las plantas , una sustancia auxética tenderá a aumentar el crecimiento celular sin ninguna división celular . Las auxinas son hormonas vegetales auxéticas .
Las auxinas ( plural de auxina / ɔː k s ɪ n / ) son una clase de hormonas vegetales (o reguladores del crecimiento de las plantas) con algunas morfógeno -como características. Las auxinas desempeñan un papel fundamental en la coordinación de muchos procesos de crecimiento y comportamiento en los ciclos de vida de las plantas y son esenciales para el desarrollo del cuerpo de la planta. El biólogo holandés Frits Warmolt Wentdescribió por primera vez las auxinas y su papel en el crecimiento de las plantas en la década de 1920. [4] Kenneth V. Thimann (1904-1997) se convirtió en el primero en aislar una de estas fitohormonas y en determinar su estructura química como ácido indol-3-acético.(IAA). Went and Thimann fue coautor de un libro sobre hormonas vegetales, Phytohormones , en 1937.
Auxinas nativas

El ácido indol-3-acético (IAA) es la auxina más abundante y básica que se produce y funciona de forma nativa en las plantas. Genera la mayoría de los efectos de la auxina en plantas intactas, y es la auxina nativa más potente.
Hay cuatro auxinas más sintetizadas endógenamente en las plantas. [1] [2]
Todas las auxinas son compuestos con anillo aromático y un grupo ácido carboxílico: [2] [3]
Hay cuatro auxinas más sintetizadas endógenamente en las plantas. [1] [2]
Todas las auxinas son compuestos con anillo aromático y un grupo ácido carboxílico: [2] [3]

Ácido 4-cloroindol-3-acético (4-CI-IAA)


Ácido indol-3-butírico (IBA)

Ácido indol-3-propiónico (IPA)
Descripción general [ editar ]
Las auxinas fueron las primeras hormonas vegetales que se descubrieron. Derivan su nombre de la palabra griega αυξειν ( auxein - "crecer / aumentar"). La auxina (es decir, IAA) está presente en todas las partes de una planta, aunque en concentraciones muy diferentes. La concentración en cada posición es información crucial para el desarrollo, por lo que está sujeta a una regulación estricta a través del metabolismo y el transporte. El resultado es que la auxina crea "patrones" de concentración de auxina máxima y mínima en el cuerpo de la planta, que a su vez guían el desarrollo adicional de las células respectivas y, en última instancia, de la planta en su conjunto.
El patrón (dinámico y sensible al medio ambiente) de la distribución de auxinas dentro de la planta es un factor clave para el crecimiento de la planta, su reacción a su entorno, y específicamente para el desarrollo de los órganos de la planta [5] [6] (como hojas o flores ). Se logra a través del transporte activomuy complejo y bien coordinado de moléculas de auxina de célula a célula por todo el cuerpo de la planta, mediante el llamado transporte de auxina polar . [5]Por lo tanto, una planta puede (en conjunto) reaccionar a condiciones externas y adaptarse a ellas, sin requerir un sistema nervioso.. Las auxinas suelen actuar en concierto con, o en oposición a, otras hormonas vegetales. Por ejemplo, la proporción de auxina a citoquinina en ciertos tejidos vegetales determina el inicio de la raíz frente a las yemas de brotes.
A nivel molecular, todas las auxinas son compuestos con un anillo aromático y un grupo ácido carboxílico . [3] El miembro más importante de la familia de las auxinas es el ácido indol-3-acético (IAA), [7] que genera la mayoría de los efectos de las auxinas en plantas intactas, y es la auxina nativa más potente. Y como auxina nativa, su estabilidad se controla de muchas maneras en las plantas, desde la síntesis, hasta la posible conjugación y la degradación de sus moléculas, siempre de acuerdo con los requisitos de la situación.
- Cinco de origen natural (endógenas) auxinas en plantas incluyen el ácido indol-3-acético, ácido 4-cloroindol-3-acético , ácido fenilacético , ácido indol-3-butírico , y ácido indol-3-propiónico . [1] [2] Sin embargo, la mayoría de los conocimientos descritos hasta ahora en biología de la auxina y como se describe en el artículo a continuación, se aplican básicamente a IAA; Las otras tres auxinas endógenas parecen tener una importancia marginal para las plantas intactas en ambientes naturales. Junto con las auxinas endógenas, los científicos y los fabricantes han desarrollado muchos compuestos sintéticos con actividad auxínica.
- Los análogos de auxina sintéticos incluyen ácido 1-naftalenacético , ácido 2,4-diclorofenoxiacético (2,4-D), [1]y muchos otros.
Algunas auxinas sintéticas, como el 2,4-D y el ácido 2,4,5-triclorofenoxiacético (2,4,5-T), también se utilizan como herbicidas . Las plantas de hoja ancha ( dicots ), como los dientes de león , son mucho más susceptibles a las auxinas que las plantas de hoja estrecha ( monocots ), como los pastos y los cultivos de cereales , por lo que estas auxinas sintéticas son valiosas como herbicidas sintéticos.
Descubrimiento de la auxina [ editar ]
Charles Darwin [ editar ]
En 1881, Charles Darwin y su hijo Francis realizaron experimentos con coleoptiles , las vainas encerrando hojas jóvenes en germinación de plántulas. El experimento expuso el coleoptil a la luz de una fuente unidireccional, y observó que se inclinaban hacia la luz. [8] Al cubrir varias partes de los coleóptiles con una tapa opaca impermeable a la luz, los Darwin descubrieron que la punta del coleoptil detecta la luz, pero que la flexión se produce en el hipocotilo.. Sin embargo, las plántulas no mostraron signos de desarrollo hacia la luz si la punta estaba cubierta con una tapa opaca, o si la punta fue removida. Los Darwin llegaron a la conclusión de que la punta del coleoptil era responsable de detectar la luz, y propusieron que un mensajero se transmite hacia abajo desde la punta del coleoptil, lo que hace que se doble. [9]
Peter Boysen-Jensen [ editar ]
En 1913, el científico danés Peter Boysen-Jensen demostró que la señal no estaba paralizada sino que era móvil. Separó la punta del resto del coleoptilo por un cubo de gelatina que impidió el contacto celular pero permitió que pasaran los químicos. Las plántulas respondían normalmente inclinándose hacia la luz. Sin embargo, cuando la punta estaba separada por una sustancia impermeable, no había curvatura del vástago. [10]
Frits fue [ editar ]
En 1928, el botánico holandés Frits Warmolt Went mostró que un mensajero químico se difunde desde las puntas de los coleoptiles. El experimento de Went identificó cómo una sustancia química que promueve el crecimiento hace que un coleoptil crezca hacia la luz. Cortó las puntas de los coleóptiles y los colocó en la oscuridad, colocando algunas puntas en bloques de agar que predijo que absorberían el químico que promueve el crecimiento. En el control de coleoptiles, colocó un bloque que carecía del químico. En otros, colocó bloques que contenían el químico, ya sea centrado en la parte superior del coleoptil para distribuir el químico uniformemente o compensado para aumentar la concentración en un lado. [9]
Cuando el producto químico promotor del crecimiento se distribuyó uniformemente, el coleoptil creció recto. Si el producto químico se distribuía de forma desigual, el coleoptil se curvaba hacia un lado del cubo, como si creciera hacia la luz, aunque se cultivara en la oscuridad. Más tarde, se propuso que la sustancia mensajera es una hormona promotora del crecimiento, a la que llamó auxina, que se distribuye de forma asimétrica en la región de flexión. Llegamos a la conclusión de que la auxina se encuentra en una mayor concentración en el lado sombreado, lo que promueve el alargamiento de las células, lo que hace que los coleóptiles se inclinen hacia la luz. [10]
Actividad hormonal [ editar ]
Las auxinas ayudan al desarrollo en todos los niveles en las plantas, desde el nivel celular , a través de los órganos y, en última instancia, a toda la planta.
Mecanismos moleculares [ editar ]
Cuando una célula vegetal entra en contacto con la auxina, causa cambios dramáticos en la expresión génica , con muchos genes regulados hacia arriba o hacia abajo. Los mecanismos precisos por los cuales esto ocurre son todavía un área de investigación activa, pero ahora hay un consenso general sobre al menos dos vías de señalización de auxinas. [11] [12]
Percepción [ editar ]
Los receptores de auxina mejor caracterizados son la familia de proteínas F-box TIR1 / AFB . Las proteínas de la caja F se dirigen a otras proteínas para su degradación a través de la vía de degradación de la ubiquitina . Cuando las proteínas TIR1 / AFB se unen a la auxina, la auxina actúa como un "pegamento molecular" que permite que estas proteínas se unan a sus objetivos (ver más abajo).
Otra proteína de unión a la auxina, ABP1 ahora se considera a menudo como un receptor de auxina (en el apoplasto ), pero en general se considera que tiene un papel mucho menor que la vía de señalización TIR1 / AFB, y mucho menos se sabe acerca de la señalización ABP1. [12]
Módulos de señalización Aux / IAA y ARF [ editar ]

Los factores de respuesta a la auxina (ARF) son un gran grupo de factores de transcripción que actúan en la señalización de la auxina. En ausencia de auxina, los ARF se unen a una clase de represores conocidos como Aux / IAA. Aux / IAA suprime la capacidad de los ARF para mejorar la transcripción de genes. Además, la unión de Aux / IAA a los ARF pone a Aux / IAA en contacto con los promotores de los genes regulados por auxina. Cuando se encuentran en estos promotores, Aux / IAA reprime la expresión de estos genes a través del reclutamiento de otros factores para realizar modificaciones en la estructura del ADN .
La unión de la auxina a TIR1 / AFB les permite unirse a las Aux / IAA. Cuando están vinculados por TIR1 / AFB, los Aux / IAA están marcados para degradación. La degradación de Aux / IAA libera proteínas ARF, que luego son capaces de activar o reprimir genes en cuyos promotores están unidos. [11] [12]
Se cree que la gran cantidad de pares de unión Aux / IAA y ARF, y que existen diferentes distribuciones entre los tipos de células y en la edad de desarrollo, explica las asombrosamente diversas respuestas que produce la auxina.
En junio de 2018 se demostró que los tejidos de las plantas pueden responder a la auxina de una manera dependiente de TIR1 de manera extremadamente rápida (probablemente demasiado rápido para ser explicado por los cambios en la expresión génica). Esto ha llevado a algunos científicos a sugerir que existe una vía de señalización de auxina dependiente de TIR1 aún no identificada que difiere de la respuesta transcripcional bien conocida. [13]
A nivel celular [ editar ]
A nivel celular, la auxina es esencial para el crecimiento celular , afectando tanto la división celular como la expansión celular. El nivel de concentración de auxina, junto con otros factores locales, contribuye a la diferenciación celular y la especificación del destino celular.
Dependiendo del tejido específico, la auxina puede promover el alargamiento axial (como en los brotes), la expansión lateral (como en el hinchamiento de la raíz) o la expansión isodiamétrica (como en el crecimiento de la fruta). En algunos casos (crecimiento de coleoptil), la expansión celular promovida por auxina ocurre en ausencia de división celular. En otros casos, la división celular promovida por auxina y la expansión celular pueden secuenciarse estrechamente dentro del mismo tejido (iniciación de la raíz, crecimiento de la fruta). En una planta viva, las auxinas y otras hormonas vegetales casi siempre parecen interactuar para determinar los patrones de desarrollo de la planta.
Patrones de organos [ editar ]
El crecimiento y la división de las células de la planta dan como resultado el crecimiento del tejido , y el crecimiento específico del tejido contribuye al desarrollo de los órganos de la planta .
El crecimiento de las células contribuye al tamaño de la planta, el crecimiento localizado de manera desigual produce la flexión, el giro y la direccionalización de los órganos; por ejemplo, los tallos giran hacia fuentes de luz ( fototropismo ), las raíces crecen en respuesta a la gravedad ( gravitropismo ) y otros tropismos se originan porque las células un lado crece más rápido que las células en el otro lado del órgano. Por lo tanto, el control preciso de la distribución de auxinas entre diferentes células tiene una importancia primordial para la forma resultante de crecimiento y organización de las plantas.
Transporte de auxina y la distribución desigual de auxina [ editar ]
Para causar el crecimiento en los dominios requeridos, las auxinas deben necesariamente estar activas preferentemente en ellas. Las auxinas máximas locales pueden formarse por biosíntesis activa en ciertas células de los tejidos, por ejemplo, a través de vías dependientes de triptófano, [14] pero las auxinas no se sintetizan en todas las células (incluso si las células conservan la capacidad potencial de hacerlo, solo en condiciones específicas ¿Se activará en ellos la síntesis de auxinas? Para ese propósito, las auxinas no solo deben ser trasladadas a los sitios donde se necesitan, sino que también deben tener un mecanismo establecido para detectar esos sitios. Para ese propósito, las auxinas deben ser translocadas hacia aquellos sitios donde se necesitan. La translocación se realiza en todo el cuerpo de la planta, principalmente desde los picos de broteshasta los picos de las raíces (de arriba a abajo).
Para distancias largas, la reubicación se produce a través del flujo de fluido en los vasos del floema , pero, para el transporte a corta distancia, se explota un sistema único de transporte polar coordinado directamente de celda a celda. Este transporte activo de corta distancia exhibe algunas propiedades morfogenéticas .
Este proceso, el transporte de auxinas polares , es direccional, está muy estrictamente regulado y se basa en una distribución desigual de los transportadores de salida de auxinas en la membrana plasmática, que envía las auxinas en la dirección correcta. Si bien las proteínas FORMADAS CON PIN (PIN) son vitales en el transporte de auxina de manera polar, [6] [15] la familia de genes AUXIN1 / LIKE-AUX1 (AUX / LAX) codifica los portadores de afluencia de auxina no polares. [dieciséis]
La regulación de la localización de la proteína PIN en una célula determina la dirección del transporte de auxina desde la célula, y el esfuerzo concentrado de muchas células crea picos de auxina o auxina máxima (regiones que tienen células con mayor auxina, un máximo). [6] La auxina máxima adecuada y oportuna dentro de las raíces y brotes en desarrollo es necesaria para organizar el desarrollo del órgano. [5] [17] [18] Máximos de auxinas circundantes son celdas con bajos niveles de auxinas, o mínimos de auxinas. Por ejemplo, en la fruta de Arabidopsis , se ha demostrado que los mínimos de auxina son importantes para el desarrollo de sus tejidos. [19]
La auxina tiene un efecto significativo en las expresiones genéticas espaciales y temporales durante el crecimiento de los meristemas apicales. Estas interacciones dependen tanto de la concentración de Auxina como de la orientación espacial durante el posicionamiento primordial. Auxin se basa en PIN1, que funciona como un portador de eflujo auxina. El posicionamiento de PIN1 sobre las membranas determina el flujo direccional de la hormona de concentraciones más altas a más bajas. [20] La iniciación de primordios en los meristemas apicales se correlaciona con niveles elevados de auxina. [21] Los genes requeridos para especificar la identidad de las células se ordenan y expresan en función de los niveles de auxina. STM (SHOOTMERISTEMLESS), que ayuda a mantener las células no diferenciadas, se regula a la baja en presencia de auxina. Esto permite que las células en crecimiento se diferencien en varios tejidos vegetales. Los genes CUC (CUP-SHAPED COTYLEDON) establecen los límites para los tejidos en crecimiento y promueven el crecimiento. [22] Se regulan al alza a través de la afluencia de auxinas. [23] Los experimentos que utilizan la visualización de GFP (PROTEÍNA DE FLUORESCENCIA VERDE) en Arabidopsis han apoyado estas afirmaciones.
Organización de la planta [ editar ]
Como las auxinas contribuyen a la conformación de los órganos, [5] [6] también son fundamentalmente necesarias para el correcto desarrollo de la propia planta. [5] Sin la regulación y organización hormonales, las plantas serían simplemente un montón de células similares en proliferación. El empleo de auxina comienza en el embrión de la planta, donde la distribución direccional de la auxina marca el crecimiento y desarrollo posterior de los polos primarios de crecimiento, y luego forma los brotes de los órganos futuros. A continuación, ayuda a coordinar el desarrollo adecuado de los órganos que surgen, como las raíces, los cotiledones y las hojas, y media las señales de larga distancia entre ellos, contribuyendo así a la arquitectura general de la planta. [5] A lo largo de la vida de la planta, la auxina ayuda a la planta a mantener la polaridad del crecimiento, [5]y en realidad "reconoce" donde tiene sus ramas (o cualquier órgano) conectadas.
Un principio importante de la organización de la planta basada en la distribución de auxinas es la dominancia apical , que significa que la auxina producida por el brote apical (o punta de crecimiento) se difunde (y se transporta) hacia abajo e inhibe el desarrollo de un crecimiento posterior del brote lateral, que de otro modo competiría con La punta apical para luz y nutrientes. La eliminación de la punta apical y su auxina de acción supresora permite que se desarrollen los brotes laterales latentes más bajos, y los brotes entre el tallo de la hoja y el tallo producen nuevos brotes que compiten para convertirse en el crecimiento principal. El proceso es bastante complejo porque la auxina transportada hacia abajo desde la punta del brote del plomo tiene que interactuar con otras hormonas vegetales (como las estrigolactonas o las citoquininas).) en el proceso en varias posiciones a lo largo del eje de crecimiento en el cuerpo de la planta para lograr este fenómeno. Este comportamiento de la planta se utiliza en la poda por los horticultores.
Finalmente, la suma de auxinas que llegan de los tallos a las raíces influye en el grado de crecimiento de las raíces. Si se eliminan las puntas de los brotes, la planta no reacciona solo por el crecimiento de los brotes laterales, que se supone que reemplazan al plomo original. También se deduce que una cantidad más pequeña de auxina que llega a las raíces da como resultado un crecimiento más lento de las raíces y los nutrientes se invierten posteriormente en mayor grado en la parte superior de la planta, que por lo tanto comienza a crecer más rápido.
Efectos [ editar ]


Auxin participa en fototropismo , geotropismo , hidrotropismo y otros cambios en el desarrollo. La distribución desigual de la auxina, debido a señales ambientales, como la luz unidireccional o la fuerza de gravedad, da como resultado un crecimiento desigual del tejido de la planta y, en general, la auxina gobierna la forma y la forma del cuerpo de la planta, la dirección y la fuerza de crecimiento de todos los órganos, y su interacción mutua. [6]
La auxina estimula la elongación celular al estimular los factores de aflojamiento de la pared, como las elastinas , para aflojar las paredes celulares . El efecto es más fuerte si las giberelinas también están presentes. La auxina también estimula la división celular si hay citoquininas presentes. Cuando se aplican auxina y citoquinina al callo , se puede generar enraizamiento si la concentración de auxina es más alta que la concentración de citoquinina. Los tejidos de xilema pueden generarse cuando la concentración de auxina es igual a las citoquininas.
La auxina también induce la acumulación de azúcar y minerales en el sitio de aplicación.
Respuesta a la herida [ editar ]
La auxina induce la formación y organización de floema y xilema . Cuando la planta está herida, la auxina puede inducir la diferenciación celular y la regeneración de los tejidos vasculares. [24]
Crecimiento de las raíces y el desarrollo [ editar ]
Las auxinas promueven la iniciación de la raíz. [25] La auxina induce tanto el crecimiento de raíces preexistentes como la formación de raíces adventicias, es decir, la ramificación de las raíces. A medida que más auxina nativa se transporta desde el tallo a las raíces, se estimula el desarrollo general de las raíces. Si se elimina la fuente de auxina, por ejemplo, al recortar las puntas de los tallos, las raíces se estimulan menos en consecuencia, y el crecimiento del tallo es compatible.
En la horticultura, las auxinas, especialmente NAA y IBA , se aplican comúnmente para estimular la iniciación de la raíz cuando se enraizan los esquejes de las plantas. Sin embargo, las altas concentraciones de auxina inhiben la elongación de la raíz y, en cambio, mejoran la formación adventicia de la raíz. La eliminación de la punta de la raíz puede conducir a la inhibición de la formación de la raíz secundaria.
La dominancia apical [ editar ]
La auxina induce la dominación apical del brote ; las yemas axilares son inhibidas por la auxina, ya que una alta concentración de auxina estimula directamente la síntesis de etileno en las yemas axilares, causando la inhibición de su crecimiento y la potenciación de la dominancia apical. Cuando se elimina el ápice de la planta, se elimina el efecto inhibitorio y se mejora el crecimiento de las yemas laterales. La auxina se envía a la parte de la planta que está alejada de la luz, donde promueve el alargamiento de las células, lo que hace que la planta se doble hacia la luz. [26]
El crecimiento del fruto y desarrollo [ editar ]
La auxina es necesaria para el crecimiento y desarrollo de la fruta y retrasa la senescencia de la fruta . Cuando se retiran las semillas de las fresas, se detiene el crecimiento de la fruta; La auxina exógena estimula el crecimiento de los frutos con las semillas eliminadas. Para frutas con semillas no fertilizadas, la auxina exógena produce parthenocarpy (crecimiento de "fruta virgen").
Las frutas forman morfologías anormales cuando se altera el transporte de auxinas. [27] En las frutas de Arabidopsis , la auxina controla la liberación de semillas de la fruta (vaina). Los márgenes de las válvulas son un tejido especializado en las vainas que regulan cuándo se abrirá la cápsula (dehiscencia). La auxina debe retirarse de las celdas del margen de la válvula para permitir que se formen los márgenes de la válvula. Este proceso requiere la modificación de los transportadores de auxina (proteínas PIN). [19]
Floración [ editar ]
La auxina también desempeña un papel menor en el inicio de la floración y el desarrollo de los órganos reproductivos. En bajas concentraciones, puede retrasar la senescencia de las flores. Se han descrito varios mutantes de plantas que afectan la floración y tienen deficiencias en la síntesis o el transporte de auxinas. En el maíz, un ejemplo es la inflorescencia estéril bif2 . [28]
Etileno biosíntesis [ editar ]
En bajas concentraciones, la auxina puede inhibir la formación de etileno y el transporte de precursores en plantas; sin embargo, altas concentraciones pueden inducir la síntesis de etileno. [29] Por lo tanto, la alta concentración puede inducir la feminidad de las flores en algunas especies. [ cita requerida ]
La auxina inhibe la abscisión antes de la formación de la capa de abscisión y, por lo tanto, inhibe la senescencia de las hojas.
Auxinas sintéticas [ editar ]
En el curso de la investigación sobre la biología de las auxinas, se sintetizaron muchos compuestos con una actividad de auxinas notable. Se ha encontrado que muchos de ellos tienen un potencial económico para el crecimiento controlado por el hombre y el desarrollo de plantas en agronomía. Las auxinas sintéticas incluyen los siguientes compuestos:
- Galería de auxinas sintéticas.
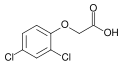

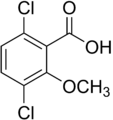







Las auxinas son tóxicas para las plantas en grandes concentraciones; Son más tóxicos para dicots y menos para monocots . Debido a esta propiedad, se han desarrollado y utilizado herbicidas de auxina sintética , incluidos 2,4-D y 2,4,5-T , para el control de malezas .
Sin embargo, algunas auxinas sintetizadas de forma exógena, especialmente el ácido 1-naftalenoacético (NAA) y el ácido indol-3-butírico (IBA), también se aplican comúnmente para estimular el crecimiento de la raíz cuando se realizan esquejes de plantas o para diferentes fines agrícolas, como la prevención de la fruta. caer en los huertos.
Usada en altas dosis, la auxina estimula la producción de etileno . El exceso de etileno (también hormona vegetal nativa) puede inhibir el crecimiento de la elongación, hacer que las hojas caigan ( abscisión ) e incluso matar la planta. Algunas auxinas sintéticas, como el 2,4-D y el ácido 2,4,5-triclorofenoxiacético (2,4,5-T) también se comercializaron como herbicidas . Los dicots , como los dientes de león , son mucho más susceptibles a las auxinas que los monocots , como los pastos y los cereales . Por lo tanto, estas auxinas sintéticas son valiosas como herbicidas sintéticos. El 2,4-D fue el primer herbicida ampliamente usado, y aún lo es. [30] El 2,4-D fue comercializado por primera vez por la compañía Sherwin-Williams y se usó a fines de la década de 1940. Es fácil y barato de fabricar.
También se ha demostrado que el 3,5,6-TPA, conocido como herbicida, aumenta el tamaño de la fruta en las plantas. A concentraciones elevadas, la hormona puede ser letal. Se ha demostrado que la dosificación hasta la concentración correcta altera las vías fotosintéticas. Este obstáculo para la planta provoca una respuesta que aumenta la producción de carbohidratos y conduce a cuerpos de fructificación más grandes. [31]
- Fabricación de herbicidas
El defoliante Agent Orange , utilizado extensivamente por las fuerzas británicas en la Emergencia Malaya y las fuerzas estadounidenses en la Guerra de Vietnam , fue una mezcla de 2,4-D y 2,4,5-T. El compuesto 2,4-D todavía está en uso y se cree que es seguro, pero la Agencia de Protección Ambiental de EE. UU. Prohibió más o menos el 2,4,5-T en 1979. El DCDE de dioxinas es un contaminante inevitable que se produce en el Fabricación de 2,4,5-T. Como resultado de la contaminación por dioxinas integrales, el 2,4,5-T se ha relacionado con leucemia , abortos espontáneos , defectos de nacimiento , daño hepático y otras enfermedades .
No hay comentarios:
Publicar un comentario