Función y química.
Cloroplastos de células de guarda
A diferencia de la mayoría de las células epidérmicas, las células protectoras de los estomas vegetales contienen cloroplastos relativamente bien desarrollados. [136] Sin embargo, exactamente lo que hacen es controvertido. [141]
Planta de inmunidad innata
Las plantas carecen de células inmunitarias especializadas: todas las células vegetales participan en la respuesta inmunitaria de la planta . Los cloroplastos, junto con el núcleo , la membrana celular y el retículo endoplásmico , [142] son actores clave en la defensa contra patógenos . Debido a su papel en la respuesta inmune de una célula vegetal, los patógenos frecuentemente se dirigen al cloroplasto. [142]
Las plantas tienen dos respuestas inmunitarias principales: la respuesta hipersensible , en la que las células infectadas se sellan y sufren la muerte celular programada , y la resistencia sistémica adquirida , donde las células infectadas emiten señales que advierten al resto de la planta de la presencia de un patógeno. Los cloroplastos estimulan ambas respuestas dañando deliberadamente su sistema fotosintético, produciendo especies reactivas de oxígeno . Altos niveles de especies reactivas de oxígeno causarán la respuesta hipersensible . Las especies reactivas de oxígeno también matan directamente cualquier agente patógeno dentro de la célula. Niveles más bajos de especies reactivas de oxígeno inician la resistencia sistémica adquirida, desencadenando la producción de moléculas de defensa en el resto de la planta. [142]
En algunas plantas, se sabe que los cloroplastos se mueven más cerca del sitio de la infección y del núcleodurante una infección. [142]
Los cloroplastos pueden servir como sensores celulares. Después de detectar el estrés en una célula, que podría deberse a un patógeno, los cloroplastos comienzan a producir moléculas como el ácido salicílico , el ácido jasmónico , el óxido nítrico y las especies reactivas de oxígeno que pueden servir como señales de defensa. Como señales celulares, las especies reactivas de oxígeno son moléculas inestables, por lo que probablemente no abandonen el cloroplasto, sino que transmiten su señal a una segunda molécula mensajera desconocida. Todas estas moléculas inician la señalización retrógrada: señales del cloroplasto que regulan la expresión génicaen el núcleo. [142]
Además de la señalización de defensa, los cloroplastos, con la ayuda de los peroxisomas , [143] ayudan a sintetizar una importante molécula de defensa, el jasmonato . Los cloroplastos sintetizan todos los ácidos grasosen una célula vegetal [142] [144] : el ácido linoleico , un ácido graso, es un precursor del jasmonato. [142]
Fotosíntesis
Una de las funciones principales del cloroplasto es su papel en la fotosíntesis , el proceso mediante el cual la luz se transforma en energía química, para producir posteriormente alimentos en forma de azúcares . El agua (H 2O) y el dióxido de carbono (CO 2 ) se utilizan en la fotosíntesis, y el azúcar y el oxígeno (O 2 ) se producen utilizando energía de la luz . La fotosíntesis se divide en dos etapas: las reacciones a la luz , donde el agua se divide para producir oxígeno, y las reacciones oscuras , o el ciclo de Calvin, que construye moléculas de azúcar a partir de dióxido de carbono. Las dos fases están unidas por los portadores de energía adenosina trifosfato (ATP) y nicotinamida adenina dinucleótido fosfato (NADP + ). [145] [146]
Reacciones de luz

Las reacciones a la luz tienen lugar en las membranas tilacoides. Toman la energía de la luz y la almacenan en NADPH , una forma de NADP + y ATPpara alimentar las reacciones oscuras.
Portadores de energía
ATP es la versión fosforilada del difosfato de adenosina (ADP), que almacena energía en una célula y potencia la mayoría de las actividades celulares. ATP es la forma activada, mientras que ADP es la forma agotada (parcialmente). NADP + es un portador de electrones que transporta electrones de alta energía. En las reacciones a la luz, se reduce , lo que significa que recoge electrones, convirtiéndose en NADPH .
Fotofosforilacion
Al igual que las mitocondrias, los cloroplastos utilizan la energía potencial almacenada en un H + , o gradiente de iones de hidrógeno para generar energía ATP. Los dos fotosistemas capturan la energía de la luz para energizar los electrones tomados del agua y liberarlos a través de una cadena de transporte de electrones . Las moléculasentre los fotosistemas aprovechan la energía de los electrones para bombear iones de hidrógeno al espacio tilacoideo, creando un gradiente de concentración , con más iones de hidrógeno (hasta mil veces más) [98] dentro del sistema tilacoide que en el estroma. Los iones de hidrógeno en el espacio tilacoideo se difundenretroceda su gradiente de concentración, fluyendo hacia el estroma a través de la ATP sintasa . La ATP sintasa utiliza la energía de los iones de hidrógeno que fluyen para fosforilar el difosfato de adenosina en trifosfato de adenosina , o ATP. [98] [147] Debido a que la ATP sintasa de cloroplasto se proyecta hacia el estroma, la ATP se sintetiza allí, en posición para usarse en las reacciones oscuras. [148]
Reducción de NADP +
Los electrones a menudo se eliminan de las cadenas de transporte de electrones para cargar NADP + con electrones, reduciéndolos a NADPH . Al igual que la ATP sintasa, la ferredoxina-NADP + reductasa , la enzima que reduce el NADP + , libera el NADPH que produce en el estroma, justo donde es necesario para las reacciones oscuras. [148]
Debido a que la reducción de NADP + elimina los electrones de las cadenas de transporte de electrones, deben reemplazarse: el trabajo del fotosistema II , que divide las moléculas de agua (H 2 O) para obtener los electrones de sus átomos de hidrógeno . [98] [145]
Fotofosforilación cíclica
Mientras que el fotosistema II fotoliza el agua para obtener y energizar nuevos electrones, el fotosistemasimplemente vuelve a energizar los electrones agotados al final de una cadena de transporte de electrones. Normalmente, los electrones reactivados son tomados por NADP + , aunque a veces pueden fluir hacia atrás por más cadenas de transporte de electrones de bombeo de H + para transportar más iones de hidrógeno al espacio tilacoide para generar más ATP. Esto se denomina fotofosforilación cíclica porque los electrones se reciclan. La fotofosforilación cíclica es común en las plantas C 4 , que necesitan más ATP que NADPH . [132]
Reacciones oscuras

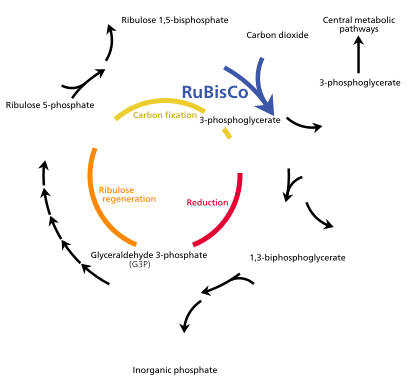


El ciclo de Calvin , también conocido como reacciones oscuras , es una serie de reacciones bioquímicas que fijan el CO 2 en moléculas de azúcar G3P y utilizan la energía y los electrones del ATP y NADPH producidos en las reacciones de luz. El ciclo de Calvin tiene lugar en el estroma del cloroplasto. [132]
Si bien se denominan "reacciones oscuras" , en la mayoría de las plantas, tienen lugar en la luz, ya que las reacciones oscuras dependen de los productos de las reacciones luminosas. [13]
Fijación de carbono y síntesis de G3P.
El ciclo de Calvin comienza utilizando la enzima RuBisCO para fijar el CO 2 en moléculas de bifosfato de ribulosa de cinco carbonos (RuBP). El resultado son moléculas de seis carbonos inestables que se descomponen inmediatamente en moléculas de tres carbonos llamadas ácido 3-fosfoglicérico o 3-PGA. El ATP y el NADPHrealizados en las reacciones de luz se utilizan para convertir la 3-PGA en gliceraldehído-3-fosfato , o moléculas de azúcar G3P. La mayoría de las moléculas de G3P se reciclan de nuevo en RuBP utilizando energía de más ATP, pero una de cada seis producidas deja el ciclo: el producto final de las reacciones oscuras. [132]
Azúcares y almidones
El gliceraldehído-3-fosfato se puede duplicar para formar moléculas de azúcar más grandes como la glucosa y la fructosa . Estas moléculas se procesan y, a partir de ellas, se produce la sacarosa aún más grande , un disacárido comúnmente conocido como azúcar de mesa, aunque este proceso tiene lugar fuera del cloroplasto, en el citoplasma . [149]

Alternativamente, los monómeros de glucosa en el cloroplasto pueden unirse entre sí para formar almidón , que se acumula en los granos de almidón que seencuentran en el cloroplasto. [149] Bajo condiciones tales como altas concentraciones de CO 2 en la atmósfera , estos granos de almidón pueden crecer mucho, distorsionando la grana y los tilacoides. Los gránulos de almidón desplazan a los tilacoides, pero los dejan intactos. [150] Las raícesanegadas también pueden causar la acumulación de almidón en los cloroplastos, posiblemente debido a que se exporta menos sacarosa fuera del cloroplasto (o más precisamente, la célula vegetal ). Esto agota una plantaSuministro de fosfato libre , que estimula indirectamente la síntesis de almidón cloroplástico. [150] Si bien están vinculados a bajas tasas de fotosíntesis, los granos de almidón en sí mismos no necesariamente interfieren significativamente con la eficiencia de la fotosíntesis, [151] y pueden ser simplemente un efecto secundario de otro factor depresor de la fotosíntesis.
Fotorrespiración
La fotorrespiración puede ocurrir cuando la concentración de oxígeno es demasiado alta. RuBisCO no puede distinguir muy bien entre oxígeno y dióxido de carbono, por lo que accidentalmente puede agregar O 2 en lugar de CO 2 a RuBP . Este proceso reduce la eficiencia de la fotosíntesis: consume ATP y oxígeno, libera CO 2 y no produce azúcar. Puede desperdiciar hasta la mitad del carbono fijado por el ciclo de Calvin. [145] Varios mecanismos han evolucionado en diferentes linajes que elevan la concentración de dióxido de carbono en relación con el oxígeno dentro del cloroplasto, aumentando la eficiencia de la fotosíntesis. Estos mecanismos se denominan mecanismos de concentración de dióxido de carbono , o MCP. Éstos incluyenMetabolismo del ácido crassulaceano , fijación de carbono C 4 , [145] y pirenoides . Los cloroplastos en plantas C 4 son notables, ya que exhiben un dimorfismo de cloroplasto distinto .
pH
Debido al gradiente de H + a través de la membrana tilacoide, el interior del tilacoide es ácido , con un pHalrededor de 4, [152] mientras que el estroma es ligeramente básico, con un pH de alrededor de 8. [153] El pH óptimo del estroma para el ciclo de Calvin es 8.1, y la reacción casi se detiene cuando el pH cae por debajo de 7.3. [154]
El CO 2 en el agua puede formar ácido carbónico , que puede alterar el pH de los cloroplastos aislados, lo que interfiere con la fotosíntesis, aunque el CO 2 se utiliza en la fotosíntesis. Sin embargo, los cloroplastos en las células vegetales vivas no se ven tan afectados por esto. [153]
Los cloroplastos pueden bombear iones de K + y H + dentro y fuera de ellos mismos mediante un sistema de transporte impulsado por la luz poco conocido. [153]
En presencia de luz, el pH de la luz de los tilacoides puede disminuir hasta 1,5 unidades de pH, mientras que el pH del estroma puede aumentar en casi una unidad de pH. [154]
Síntesis de aminoácidos
Los cloroplastos solos producen casi todos los aminoácidos de una célula vegetal en su estroma [155], excepto los que contienen azufre , como la cisteína y la metionina . [156] [157] La cisteína se produce en el cloroplasto (el proplastido también), pero también se sintetiza en el citosol y las mitocondrias , probablemente porque tiene problemas para cruzar las membranas para llegar a donde se necesita. [157] Se sabe que el cloroplasto produce los precursores de la metionina, pero no está claro si el orgánulo realiza la última etapa de la vía o si ocurre en el citosol.. [158]
Otros compuestos nitrogenados
Los cloroplastos producen todas las purinas y pirimidinas de una célula , las bases nitrogenadas que seencuentran en el ADN y el ARN . [155] También convierten el nitrito (NO 2 - ) en amoníaco (NH 3 ) que suministra nitrógeno a la planta para producir sus aminoácidos y nucleótidos . [155]
Otros productos quimicos
El plástido es el sitio de síntesis de lípidos diverso y complejo en las plantas. [159] [160] El carbono utilizado para formar la mayoría del lípido es del acetil-CoA , que es el producto de descarboxilación del piruvato . [159] Elpiruvato puede entrar en el plastidio del citosol por difusión pasiva a través de la membrana después de la producción en la glucólisis . [161] El piruvato también se fabrica en el plastidio a partir de fosfoenolpiruvato, un metabolito producido en el citosol a partir de piruvato o PGA . [159] El acetato en el citosol no está disponible para la biosíntesis de lípidos en el plastidio. [162]La longitud típica de los ácidos grasos producidos en el plástido son 16 o 18 carbonos, con 0-3 cis dobles enlaces. [163]
La biosíntesis de ácidos grasos a partir de acetil-CoA requiere principalmente dos enzimas. La acetil-CoA carboxilasa crea malonil-CoA, utilizada tanto en el primer paso como en los pasos de extensión de la síntesis. La ácido graso sintasa (FAS) es un gran complejo de enzimas y cofactores que incluyen la proteína transportadora de acilo (ACP) que contiene la cadena de acilo a medida que se sintetiza. El inicio de la síntesis comienza con la condensación de malonil-ACP con acetil-CoA para producir ketobutiril-ACP. 2 reducciones que involucran el uso de NADPH y una deshidratación crean butiril-ACP. La extensión del ácido graso proviene de ciclos repetidos de condensación, reducción y deshidratación de malonil-ACP. [159]
Otros lípidos se derivan de la vía del fosfato de metil-eritritol (MEP) y consisten en giberelinas , esteroles , ácido abscísico , fitol e innumerables metabolitos secundarios . [159]
Diferenciación, replicación y herencia.

Los cloroplastos son un tipo especial de orgánulo de células vegetales llamado plástido , aunque los dos términos a veces se usan indistintamente. Hay muchos otros tipos de plastidios, que llevan a cabo diversas funciones. Todos los cloroplastos en una planta descienden de proplastidos no diferenciados que se encuentran en el cigoto , [155] o huevo fertilizado. Los proplastos se encuentran comúnmente en los meristemas apicales de una planta adulta . Los cloroplastos normalmente no se desarrollan a partir de proplastos en los meristemas de la punta de la raíz [164]; en cambio, la formación de amiloplastos que almacenan almidón es más común. [155]
En los brotes , los proplastidos de los meristemas apicales de los brotes pueden convertirse gradualmente en cloroplastos en los tejidos de las hojas fotosintéticas a medida que la hoja madura, si se expone a la luz requerida. [11] Este proceso involucra invaginaciones de la membrana plástica interna, formando láminas de membrana que se proyectan hacia el estroma interno . Estas láminas de membrana luego se doblan para formar tilacoides y grana . [165]
Si los brotes de angiospermas no están expuestos a la luz requerida para la formación de cloroplastos, los proplastos pueden convertirse en una etapa de etioplasto antes de convertirse en cloroplastos. Un etioplasto es un plástido que carece de clorofila y tiene invaginaciones de la membrana interna que forman una red de tubos en su estroma, llamado cuerpo prolamelar . Mientras que los etioplastos carecen de clorofila, tienen un precursor de clorofila amarillo en existencia . [11] A los pocos minutos de la exposición a la luz, el cuerpo prolamelar comienza a reorganizarse en pilas de tilacoides y comienza a producirse clorofila. Este proceso, donde el etioplasto se convierte en un cloroplasto, lleva varias horas. [165] GimnospermasNo requieren luz para formar cloroplastos. [165]
No hay comentarios:
Publicar un comentario