Virus a ADN
Caudovirales es un orden de virus que comprende a la mayor parte de los bacteriófagos (en torno a un 95%). En el marco del esquema de la Clasificación de Baltimore se integran en el Grupo I, ya que tienen un genoma ADNbicatenario. El tamaño del genoma está comprendido entre 18 y 500 kbp. Las partículas virales tienen una forma distintiva con una cabeza icosaedral que contiene el genoma y conectada a una cola. El orden abarca una amplia gama de virus, muchos de los cuales tienen los mismos o similares genes, mientras que la secuencia de nucleótidos puede variar considerablemente, incluso entre el mismo género. Debido a su característica estructura, se cree que comparten un origen común.
Infección
Al encuentro con una bacteria huésped, la cola del virus se une a los receptores de la superficie celular bacteriana y el virus introduce su ADN en la célula mediante el uso de un mecanismo de inyección. La cola hace un agujero a través de la pared celular y de la membrana plasmática de la bacteria y el genoma pasa por la cola a la célula. Una vez dentro, los genes se expresan mediante la transcripción hecha por la maquinaria huésped, haciendo uso de losribosomas. Típicamente, el genoma se replica haciendo uso de concatémeros.
Ensamblado y maduración
Las proteínas de la cápside se reúnen para formar una protocabeza, en el que se introduce el genoma. Una vez que esto ha ocurrido, la maduración de la protocabeza se realiza por la división de las unidades de la cápside para formar una cabeza icosaedral con simetría 5. Después de la maduración, la cola se añade por una de dos maneras: o bien la cola se construye por separado y se une con la cabeza, o bien se construye directamente sobre la cabeza del fago. La cola consiste en una hélice de proteínas con simetría 6. Después de la maduración de las partículas virales, la célula es lisada usando lisinas, holinas o una combinación de ambas.
| Caudovirales | ||
|---|---|---|
 | ||
| Clasificación de los virus | ||
| Grupo: | I (Virus ADN bicatenario) | |
| Orden: | Caudovirales | |
| Familias | ||
Caudovirales
Los Caudovirales son un pedido de virus también conocidos comobacteriophages rabudo. Según el esquema de la clasificación de Baltimore,Caudovirales son el grupo I virus ya que han varado dos veces el ADN (dsDNA) genomas, que pueden estar en todas partes de 18,000 pares de bases a 500,000 pares de bases de la longitud. Las partículas del virus tienen una forma distinta; cada virion tiene una cabeza de icosohedral que contiene el genoma viral y es atada a una cola flexible por una proteína del conector. El pedido cerca una amplia gama de virus, muchos de cual conteniendo genes de secuencia nucleotide similar y función. Unos siguieron genomas bacteriophage puede variar completamente considerablemente en la secuencia nucleotide, sin embargo, hasta entre el mismo género. Debido a su estructura característica y posesión de genes potencialmente homólogos, se cree que estos bacteriophages poseen un origen común.
Hay al menos 350 especies reconocidas en este pedido.
Infección
Para encontrar una bacteria del anfitrión, la sección de la cola del virion liga a receptores en la superficie de la célula y entrega el ADN en la célula por el uso de un mecanismo parecido a injectisome (un injectisome es un nanomachine que evolucionó para la entrega de proteínas por la secreción del tipo III). La sección de la cola del virus perfora un agujero a través de la pared de la célula bacteriana y membrana plasma y el genoma transmite la cola en la célula. Una vez dentro de los genes se expresan de transcripciones hechas por la maquinaria del anfitrión, usando al anfitrión ribosomes. Típicamente, el genoma es reproducido por el uso de concatemers, en el cual los segmentos que se superponen del ADN se hacen, y luego se reúnen para formar el genoma entero.
Asamblea y maduración
Las proteínas capsid virales vienen juntos para formar una procabeza del precursor, en la cual el genoma entra. Una vez que esto ha ocurrido, la procabeza se somete a la maduración por la hendidura de subunidades capsid para formar un icosohedral phage cabeza con la simetría de 5 pliegues. Después de la maduración principal, la cola se afilia de uno de dos modos: la cola se construye por separado y se afilia con el conector, o la cola se construye directamente en la cabeza de phage. Las colas consisten en la hélice proteínas basadas con la simetría de 6 pliegues. Después de la maduración de partículas del virus, la célula es lysed por lysins, holins, o una combinación de los dos.
Taxonomía
A causa de la carencia de la homología entre el aminoácido y las secuencias del ADN de los estos virus impide éstos de acostumbrarse como marcadores taxonómicos como es común para otros organismos, las tres familias aquí se definen sobre la base de la morfología. Este esquema de la clasificación fue originado por Bradley en 1969 y se ha ampliado desde entonces.
Todos los virus en este pedido tienen icosahedral o cabezas de oblate, pero se diferencian en la longitud y las capacidades contráctiles de las sus colas. Los Myoviridae tienen colas largas que son contráctiles; Podoviridae tienen colas no contráctiles cortas; y Siphoviridae tienen colas no contráctiles largas. Siphoviridae constituyen la mayoría de los virus seguidos conocidos.
Bradley mandó a lo que se conoce ahora como Myoviridae como el tipo A, Siphonviridae como el tipo B y Podoviridae como el tipo C. También dividió sus grupos sobre la base de la morfología principal: dentro del Grupo A, A1 tienen pequeñas cabezas isométricas; A2 tienen cabezas profallecidas; y A3 han alargado a cabezas. Dentro de grupos B y C, los números de manera similar se asignaron: B1 y C1 tienen pequeñas cabezas isométricas; B2 y C2 tienen cabezas profallecidas; y B3 y C3 han alargado a cabezas.
Evolución de Bacteriophage
Bacteriophages ocurren en más de 140 géneros bacterianos o archaeal. Se levantaron repetidamente en anfitriones diferentes y hay al menos 11 líneas separadas del descenso. Más de 5100 bacteriophages se han examinado en el microscopio de electrones desde 1959. De éstos, al menos 4950 phages (el 96%) tienen colas. De phages rabudo, el 61% tiene colas largas, no contráctiles (Siphoviridae). Phages seguidos parecen ser monophyletic y son el grupo del virus conocido más viejo.
Virus que infecta las bacterias provocando su lisis. También suelen llamarse fagos; no se diferencian en su estructura
y propiedades de los otros virus, excepto en la característica de infectar específicamente las bacterias. Su morfología
se aprecia claramente con el microscopio electrónico; constan de una cabeza hexagonal con una cubierta proteica, en cuyo
interior se encuentra una espira de ácido desoxirribonucleico (DNA) y una cola delgada formada por las moléculas de tres
proteínas distintas. Cuando el bacteriófago se pone en contacto con la bacteria, una de las proteínas de la cola se fija a la
membrana, mientras que otra, de carácter enzimático, la disuelve. A continuación la espira de DNA es inyectada en el interior
del protoplasma de la bacteria. La espira de DNA tiene una estructura y función análogas a las de los cromosomas. Al
introducirse en el interior de la bacteria sustituye la "información" del DNA del material genético de la bacteria por el propio y
aprovecha todas las reacciones del protoplasma para sintetizar nuevos bacteriófagos completos de DNA y proteínas. Cuando
toda la sustancia celular se ha convertido en bacteriófagos, la membrana bacteriana estalla ( lisis) y los fagos infectan nuevas
bacterias. La investigación de los bacteriófagos fue iniciada por Twort y d'Hérelle en Escherichia coli; en un principio se creyó
que podrían actuar como antagonistas de las bacterias en las enfermedades infecciosas. Los bacteriófagos se reconocen por
dejar espacios blancos o placas en los cultivos de bacterias; su especificidad para atacar selectivamente diversas especies e
incluso cepas se utiliza para el diagnóstico bacteriológico. En la industria son perjudiciales para todos los procesos que
requieren cepas puras de bacterias. El método para combatir sus efectos es la selección de cepas bacterianas resistentes. Los
bacteriófagos de las bacterias radicícolas fijadoras de nitrógeno son muy perjudiciaies para la agricultura. Los bacteriófagos
constiruyen un importantísimo material de experimentación para investigar la estructura de los virus y del material genético y
la transducción de genes y fragmentos de cromosomas entre las bacterias.
y propiedades de los otros virus, excepto en la característica de infectar específicamente las bacterias. Su morfología
se aprecia claramente con el microscopio electrónico; constan de una cabeza hexagonal con una cubierta proteica, en cuyo
interior se encuentra una espira de ácido desoxirribonucleico (DNA) y una cola delgada formada por las moléculas de tres
proteínas distintas. Cuando el bacteriófago se pone en contacto con la bacteria, una de las proteínas de la cola se fija a la
membrana, mientras que otra, de carácter enzimático, la disuelve. A continuación la espira de DNA es inyectada en el interior
del protoplasma de la bacteria. La espira de DNA tiene una estructura y función análogas a las de los cromosomas. Al
introducirse en el interior de la bacteria sustituye la "información" del DNA del material genético de la bacteria por el propio y
aprovecha todas las reacciones del protoplasma para sintetizar nuevos bacteriófagos completos de DNA y proteínas. Cuando
toda la sustancia celular se ha convertido en bacteriófagos, la membrana bacteriana estalla ( lisis) y los fagos infectan nuevas
bacterias. La investigación de los bacteriófagos fue iniciada por Twort y d'Hérelle en Escherichia coli; en un principio se creyó
que podrían actuar como antagonistas de las bacterias en las enfermedades infecciosas. Los bacteriófagos se reconocen por
dejar espacios blancos o placas en los cultivos de bacterias; su especificidad para atacar selectivamente diversas especies e
incluso cepas se utiliza para el diagnóstico bacteriológico. En la industria son perjudiciales para todos los procesos que
requieren cepas puras de bacterias. El método para combatir sus efectos es la selección de cepas bacterianas resistentes. Los
bacteriófagos de las bacterias radicícolas fijadoras de nitrógeno son muy perjudiciaies para la agricultura. Los bacteriófagos
constiruyen un importantísimo material de experimentación para investigar la estructura de los virus y del material genético y
la transducción de genes y fragmentos de cromosomas entre las bacterias.

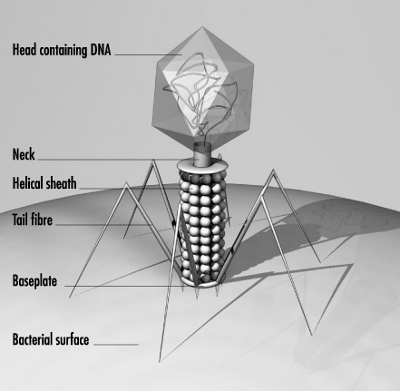
Los bacteriófagos (también llamados fagos -del griego φαγητόν (phagētón), «alimento, ingestión») son virus que infectanexclusivamente a las bacterias.
Al igual que los virus que infectan células eucariotas, los fagos están constituidos por una cubierta proteica o cápside en cuyo interior está contenido su material genético, que puede ser ADN , de 5.000 a 500.000 pares de bases. El tamaño de los fagos oscila entre 20 y 200 nm aproximadamente.
Los fagos son ubicuos y pueden ser encontrados en diversas poblaciones de bacterias, tanto en el suelo como en la flora intestinal de los animales. Uno de los ambientes más poblados por fagos y otros virus es el agua de mar, donde se estima que puede haber en torno a 109 partículas virales por mililitro, pudiendo estar infectadas por fagos el 70% de las bacterias marinas.
Historia
La historia del descubrimiento de los bacteriofagos ha sido objeto de muchos debates, los cuales incluyen una controversia sobre quién fue su descubridor. En 1913, el bacteriólogo británico Frederick Twort descubrió un agente bacteriolítico que infectaba y mataba a las bacterias, pero, no fue capaz de saber qué era exactamente dicho agente, señalando en una de sus hipótesis que, entre otras posibilidades, podría tratarse de un virus. No fue sino hasta 1917, cuando el microbiólogo canadiense Félix d'Herelle anunció el descubrimiento de "un invisible antagonista microbiano del bacilo de la disentería", al cual afirmó que se trataba de un virus al cual llamóbacteriófago. Los trabajos de d'Herelle iniciaron desde 1910 y en contraste con Twort y algunos otros científicos que habían reportado fenómenos similares, tuvo pocas dudas sobre la naturaleza del fenómeno que estaba observando y afirmó que se trataba de un virus que parasitaba a la bacteria. El nombre bacteriófago lo formó de la palabra "bacteria" y "phagein" (comer o devorar, en griego), implicando que los fagos "comen" o "devoran" a las bacterias.
Replicación

Los fagos pueden generar el ciclo lítico o el ciclo lisogénico, aunque muy pocos son capaces de llevar a cabo ambos. si se lleva a cabo la lisis, no puede llevarse a cabo la lisogenia y viceversa. En el ciclo lítico, las células hospedadoras del fago son lisadas (destruidas) tras la replicación y encapsulación de las partículas virales, de forma que los nuevos virus quedan libres para llevar a cabo una nueva infección.
Por el contrario, en el ciclo lisogénico no se produce la lisis inmediata de la célula. El genoma del fago puede integrase en el ADN cromosómico de la bacteria hospedadora, replicándose a la vez que lo hace la bacteria o bien puede mantenerse estable en forma de plásmido, replicándose de forma independiente a la replicación bacteriana. En cualquier caso, el genoma del fago se transmitirá a toda la progenie de la bacteria originalmente infectada. El fago queda así en estado de latencia hasta que las condiciones del medio se vean deterioradas: disminución de nutrientes, aumento de agentes mutagénicos, etc. En este momento, los fagos endógenos o profagos se activan y dan lugar al ciclo lítico que termina con la lisis celular.
Conversión lisogénica
En ocasiones, los profagos otorgan beneficios a la bacteria huésped mientras permanecen en estado letárgico al incorporarle nuevas funciones a su genoma; éste fenómeno se conoce las condiciones de vida de la bacteria.
Acoplamiento
Los fagos se acoplan a receptores específicos en la superficie de la bacteria, que pueden ser lipopolisacáridos, ácidos teicoicos, proteínas o incluso flagelos. Por ello, cada fago solo podrá infectar ciertas bacterias según sus receptores. Puesto que los fagos no son móviles, dependen de encuentros al azar con los receptores adecuados en solución para poder infectar una bacteria.
Parece que los bacteriófagos presentan una especie de jeringa mediante la cual introducen su material genético en el interior de la célula. Tras el reconocimiento del receptor adecuado, la cola y cuello del fago se contraen, quedando así el fago acoplado a la superficie celular. El material genético puede ser ahora introducido a través de la membrana o bien simplemente depositado sobre la superficie. No se descarta que pueda haber fagos con otros métodos diferentes para introducir su material genético en la célula.
Síntesis de proteínas y ácidos nucleicos
En un corto espacio de tiempo, que pueden llegar a ser minutos, los ribosomas bacterianos comienzan a traducir el ARNm viral a proteínas. En el caso de los fagos basados en ARN, una RNA-replicasa es sintetizada al inicio del proceso.
- Las proteínas producidas en la fase temprana y unas pocas proteínas que estaban presentes en el virión podrían modificar la RNA-polimerasa bacteriana de forma que transcriba preferentemente los ARNm virales. Todo el sistema de traducción y de replicación normal de la bacteria se ve interrumpido y es forzado a producir nuevas partículas virales.
- Posteriormente, las proteínas helper se encargarán de ensamblar las nuevas partículas virales.
- Finalmente, se sintetizan las proteínas de la fase tardía, involucradas en el proceso de la lisis celular.
Ensamblaje
En el caso del fago T4, la construcción de nuevas partículas virales es un complejo proceso que requiere la ayuda de ciertas moléculas. La cola y la cabeza o cápside del fago son construidas por separado y se ensamblan posteriormente de forma espontánea solamente en la lisis completa. Después, el ADN es empaquetado en el interior de la cápside mediante un mecanismo no muy bien conocido aún. Todo el proceso puede durar unos 15 minutos.La cabeza tiene simetría icosaédrica (un icosaedro con un prisma hexagonal intercalado. La cola es de simetría helicoidal formada por un tubo central rígido y una vaina contráctil. De la placa basal salen seis espículas basales y seis fibras caudales. En la conexión de la cabeza y la cola existe un collar. Cada una de estas estructuras está formada por diferentes proteínas.
Liberación de los fagos
Los fagos pueden ser liberados mediante lisis celular o por secreción celular. En el caso del fago T4, unos 20 minutos después de inyectar el material genético, más de 300 fagos son liberados vía lisis. La proteína que lleva a cabo la lisis es la endolisina, una enzima capaz de romper las moléculas de peptidoglicano de la pared bacteriana. Sin embargo, algunos fagos pueden quedarse en la célula como parásitos, de forma que la bacteria va secretando constantemente nuevas partículas virales. En estos casos, los viriones salen mediante procesos de exocitosis, en los que cada uno se queda con una pequeña porción de membrana bacteriana que los envuelve. Todos los nuevos fagos liberados quedan en disposición de infectar a una nueva bacteria.
Biología molecular
Los fagos cumplen un papel de gran importancia en la biología molecular al ser utilizados como vectores de clonación para insertar ADN dentro de las bacterias y obtener como resultado bibliotecas genómicas. Uno de los fagos más utilizados con este fin es el Fago λ.
Terapia fágica
La terapia fágica ha sido utilizada desde la década de 1940 en la ex Unión Soviética como una alternativa a los antibióticos para tratar infecciones bacterianas, ya que eliminar bacterias es lo que los fagos hacen mejor. El desarrollo de cepas bacterianas resistentes a múltiples drogas ha conducido a investigadores en medicina a reconsiderar a los fagos como una alternativa al uso de antibióticos. Hay una biblioteca de búsqueda de fagos específicos y sus usos terapéuticos en el Instituto Tbilisi, en la República de Georgia.
En agosto de 2006, la FDA (Food and Drug Administration) de Estados Unidos aprobó el uso de bacteriófagos en ciertas carnes con el fin de acabar con la bacteriaListeria monocytogenes. En el otoño de 2011 la Fundación Bill y Melinda Gates anuncia que financiará la investigación, llevada a cabo por el Centro Nacional de Biotecnología (España), dependiente del CSIC, del uso de bacteriófagos como sustituto de los antibióticos ante las infecciones del tracto digestivo producidas porbacterias.
Bacteriófagos modelo
A continuación se presenta una lista de los bacteriófagos más ampliamente estudiados:
- Fago λ - Lisógeno
- Fago T4 (169 a 170 kpb, 200 nm de tamaño)
- Fago T7
- Fago R17
- Fago M13 - Fagémido
- Fago MS2 (23-25 nm de tamaño)
- Fago G4
- Fago P1
- Fago P2
- Fago N4
- Fago Φ6
- Fago P22
- Fago Φ29
- Fago ΦX174
Familias de bacteriófagos
- Familia Corticoviridae
- Familia Cystoviridae
- Familia Inoviridae
- Familia Leviviridae
- Familia Microviridae
- Familia Myoviridae
- Familia Podoviridae
- Familia Plasmaviridae
- Familia Siphoviridae
- Familia Tectiviridae
Bacteriofagos
Los virus son moléculas de DNA o RNA rodeadas por una envoltura proteica que necesitan células viables para poder replicarse. Los virus utilizan la maquinaria metabólica de las células para sintetizar su material genético y proteínas de la envoltura. Existen distintos tipos de virus que pueden infectar células procariontes o células eucariontes. Los bacteriófagos o fagos son virus que se reproducen en células procariontes.El genoma de los fagos puede ser RNA simple cadena (MS2, Qß), RNA doble cadena (phi 6), DNA simple cadena (phi X174, fd, M13) o DNA doble cadena (T3, T7, lambda , T5, Mu, T2, T4). Estos ácidos nucleicos pueden contener bases inusuales que son sintetizadas por proteínas del fago. En los T-pares el genoma no contiene citosina sino 5'- hidroximetilcitosina, mientras que en otros tipos de fago alguna de las bases esta parcialmente sustituida.Bacteriófago T4
Replicación del Bacteriófago T4
Esquema del Ciclo de Replicación de un Bacteriófago T4El ciclo de replicación de un bacteriófago T4 se puede dividir esquemáticamente en distintas etapas, las que son comunes a otros virus bacterianos y eucarióticos.1. Adsorción2. Inyección del material genético viral3. Replicación del material genético viral4. Síntesis de las envolturas proteicas5. Empaquetamiento del DNA dentro de la envoltura proteica y ensamblaje de la envoltura6. Lisis y liberación de las partículas viralAdsorción: El virus se fija o adsorbe a componentes de la superficie celular que actúan como receptores específicos. La zona de adsorción del virus es complementaria al receptor celular, por lo tanto un determinado virus sólo puede infectar un número limitado de cepas celulares que contengan a un determinado receptor. La naturaleza de la zona de adsorción varía con el tipo de fago. En el T4 se localiza en el extremo de la cola, en donde se encuentran la placa basal, las espículas y las fibras de la cola.Esquema de los principales eventos en la adsorción del bacteriófago T4 a la pared celular de la bacteria E. coli.(a) el fago libre muestra las fibras y las espículas de la cola. (b)Adhesión de las fibras de la cola. (c) El fago se acerca a la pared celular y las espículas entran en contacto con la pared celular.Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal, alguna de las cuales tienen actividad enzimática y producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior.Esquema del mecanismo de penetración del material genético del fago T4 o T2 a través de la pared celular de la bacteria. (a) Las espículas del fago entran en contacto con la pared celular y la vaina se encuentra extendida. (b) La vaina de la cola se contrae y el material genético del fago penetra la pared celular; la lisozima presente en el fago digiere la porción de pared celular localizada directamente bajo la partícula viral.Replicación del material genético viral: El material genético viral que ingresa en una célula contiene bases modificadas que evitan la degradación por nucleasas bacterianas. Esta modificación consiste en la glicosilación y/o metilación de algunas determinadas bases. En el caso del fago T4 se glucosila la base 5'-hidroximetilcitosina. Para lograr una efectiva replicación del genoma viral se deben sintetizar algunas proteínas ni bien el material genético ingresa en la célula. Esta proteínas tempranas reparan el poro de la membrana citoplasmática por donde ingresó el genoma viral, degradan el DNA bacteriano lo que proporciona una fuente de precursores, evita la síntesis de RNA y proteínas bacterianas, y proporciona ribosomas para la síntesis de proteínas del fago. Además algunas de estas proteínas tempranas participan en la síntesis de las bases inusuales. La forma de replicación del genoma viral es dependiente del tipo de material genético (si es RNA o DNA, si es simple o doble cadena). En el caso del fago T4, las moléculas replicadas se aparean en los extremos y formando una molécula de DNA más larga denominada concatámero. Después una enzima corta esta larga molécula lineal en moléculas más pequeñas de igual longitud. Las moléculas de DNA del T4 tienen se caracterizan por estar permutadas circularmente (el DNA del T4 es lineal) de esta forma todas las moléculas de DNA resultantes contienen genes completos y funcionales. La enzima del T4 que corta al concatámero produce moléculas de DNA de tamaños similares pero no reconoce sitios específicos sobre la molécula, en cambio la enzima del T7 reconoce sitios específicos sobre el DNA.Síntesis de las envolturas proteicas: Las proteínas de la envoltura (cápside, vaina, fibras, etc) son proteínas tardías que se sintetizan después de iniciada la replicación del material genético. La síntesis de cada componente proteico se realiza separadamente. En el caso del fago T4, el material genético es encapsidado antes del ensamble del resto de los componentes.Ensamble: Todas las proteínas de la envoltura se ensamblan para formar una partícula viral madura capaz de infectar a otra célula cuando sea liberada.Lisis celular y liberación de las partículas virales: La lisis celular se debe a la síntesis de proteínas tardías codificadas en el genoma del fago. En el fago T4, estas proteínas son enzimas que lesionan la membrana citoplasmática y la pared celular.LisogeniaPoco tiempo después de que fueron descubiertos los fagos, se aislaron cepas bacterianas que parecían ser portadoras silenciosas de ciertos tipos de fagos. Los líquidos obtenidos a partir de cultivos de estas cepas portadoras mostraban la presencia de fagos; sin embargo, las cepas portadoras no eran sensibles a ser destruidas por el fago que portaban. Por otra parte, cepas de bacterias emparentadas con la cepa portadora resultaban ser sensibles al fago presente en las cepas portadoras. Las cepas de bacterias portadoras de fagos silenciosos o latentes fueron denominadas cepas lisogénicas. A principios de los años cincuenta se descubrió que los fagos son capaces de adsorberse a las bacterias lisogénicas, pero no se produce la subsecuente lisis de estas bacterias. Por otra parte, cuando el fago procedente de una cepa lisogénica es sembrado en una cepa de bacterias sensibles, se pueden aislar en estos cultivos colonias de bacterias que se comportan igual que las cepas lisogénicas. En 1950, André Lwoff cultivó una sola célula procedente de una cepa lisogénica de Bacillus megaterium y observó bajo el microscopio la división de esta bacteria. Posteriormente, Lwoff removió una de las células hijas junto con un poco del medio de cultivo. Este proceso fue repetido varias veces: cada vez que se dividía la célula remanente, era removida una de las células hijas y algo del medio de cultivo; las células hijas y el viejo medio de cultivo fueron sembrados en agar para determinar si daban origen a una población de bacterias lisogénicas y a la presencia de fago en el medio de cultivo. Estos experimentos mostraron que las bacterias lisogénicas pueden crecer y dividirse sin liberar fagos al medio de cultivo. Sin embargo, por alguna razón desconocida, algunos filtrados obtenidos a partir de extractos de bacterias lisogénicas mostraban la presencia de partículas virales, o sea, fagos. Lwoff razonó que en las cepas lisogénicas el fago se encuentra en forma de un precursor no infeccioso al que denominóprofago. La lisis de algunas de estas bacterias lisogénicas ocurre solamente cuando estas células han sido estimuladas para producir fagos. Lwoff y colaboradores observaron que la irradiación con luz ultravioleta (U.V.) era capaz de inducir la producción de fagos en una población de bacterias lisogénicas, mismas que eran lisadas en la medida que se incrementaba la concentración de fagos liberados al medio de cultivo. Por lo tanto, una bacteria lisogénica posee la capacidad de heredar el fago a sus descendientes, muy pocos de los cuales se lisarán en forma espontánea. Sin embargo, la mayor parte de la progenie de una bacteria lisogénica puede ser inducida a producir el fago por medio de la irradiación con U.V. o tratamiento con otros factores inductores. Se denomina temperados a los bacteriófagos capaces de existir en forma de profago en el interior de una bacteria hospedera. Después de que el profago ha sido inducido por irradiación, ocurre un breve periodo de eclipse en el cual no se puede detectar la presencia del fago dentro de la bacteria. Sin embargo, es posible detectar la aparición de proteínas y ácido nucleico específicos del fago; estos elementos serán ensamblados para formar los nuevos fagos maduros poco antes de que ocurra la lisis de la bacteria hospedera.En 1951, Esther Lederberg descubrió en forma accidental que la cepa de E. coli K12 era de tipo lisogénico. El fago latente en dicha cepa fue aislado al mezclar E. coli Kl2 con derivados no lisogénicos de esta cepa bacteriana. El fago resultante es ahora conocido como fago y representa el caso más estudiado del fenómeno de lisogenia.Las bacterias infectadas por fagos temperados continúan dividiéndose por varias generaciones y son inmunes o resistentes a ser superinfectadas por el mismo tipo de fago que albergan o por otros fagos pertenecientes a clases emparentadas con el fago temperado original. Estas bacterias contienen cuando menos una copia íntegra del genoma del fago. Las bacterias infectadas por fagos temperados portan la información genética correspondiente al fago, a través de múltiples divisiones bacterianas, o sea, el genoma del fago es replicado al mismo tiempo que ocurre la replicación del genoma bacteriano; este hecho hace posible que la bacteria original pueda heredar el fago temperado a la subsecuente progenie bacteriana.Transducción
La transducción fue cronológicamente el último sistema de transferencia genética bacteriana que se descubrió.En 1951 Joshua Lederberg y su colaborador Zinder estaban investigando en Salmonella la posible existencia de un sistema de conjugación al estilo del que se acababa de descubrir en su pariente Escherichia coli). Mezclaron dos cepas de Salmonella, cada una con un juego distinto de marcadores genéticos. (Eureka! Obtuvieron recombinantes. Descartaron que se tratara de transformación, ya que los resultados eran similares si añadían DNasa al sistema. Entonces, ¿era un fenómeno de conjugación? Realizaron el experimento del tubo en "U", con una membrana separando los dos brazos de la U, en cada uno de los cuales se colocaba una de las cepas. La membrana impide el paso de bacterias y los contactos intercelulares directos entre las dos cepas. Pues bien... seguía habiendo recombinantes. Esto descartaba, pues, que se tratara de conjugación. Se postuló que debía de existir un "agente filtrable" resistente a las nucleasas, responsable último de la transferencia genética.¿Cuál era la naturaleza exacta del misterioso agente filtrable? Por experimentos independientes se sabía que una de las dos cepas de Salmonella producía un fago (llamado P22), de tipo moderado. Con una serie de ensayos se demostró que era precisamente este fago el responsable de los recombinantes:el tratamiento de sobrenadantes de esa cepa con calor o con antisuero provocaba la inactivación tanto del fago como del agente filtrable;las cepas de Salmonella resistentes a P22 (porque no adsorben el fago) no pueden interaccionar con el agente filtrable, y por lo tanto tampoco dan recombinantes;finalmente, se comprobó que la cepa productora del agente filtrable poseía un fago moderado en forma de profago. La inducción de esta cepa lisogénica era la responsable de producir algunas partículas de fagos portadoras de material genético de la cepa de origen, que los fagos inyectaban posteriormente a las células de la cepa receptora.Así pues, se acababa de descubrir un nuevo sistema de transferencia genética entre bacterias, sistema que fue bautizado con el nombre de transducción.La transducción se puede definir como el proceso de transferencia genética desde una célula donadora a otra receptora mediatizado por partículas de bacteriófagos que contienen ADN genómico de la primera. En la transducción podemos distinguir dos etapas diferenciadas:1. Formación de la partícula fágica transductora: un trozo de material genético de la célula donadora se introduce en el interior de la cabeza de la cápsida de un fago. Las partículas transductoras son en cierta manera "subproductos" anómalos del ciclo normal del fago.2. La partícula transductora inyecta de forma habitual el ADN que porta a la célula receptora, donde este ADN puede eventualmente recombinarse y expresar su información.La transducción descubierta por Lederberg y Zinder se llama transducción generalizada.Mediante ella se puede transferir cualquier marcador del genóforo del donador, con aproximadamente la misma frecuencia relativa (de ahí el calificativo de generalizada).La transducción generalizada se produce sólo como consecuencia de infecciones líticas.El ADN del genomio de la bacteria donadora que es introducido en la partícula transductora suele ir sin acompañamiento de ADN del propio fago. Por ello, a esta peculiar partícula consistente en cápsida del fago que encierra sólo ADN genofórico de la bacteria se la denomina pseudovirión.Siguiendo con la buena racha de descubrimientos, pocos años más tarde (1956), el mismo Lederberg (esta vez junto con su mujer, y con Morse) hallaron un tipo nuevo de transducción, mientras estaban estudiando el sistema del fago moderado l y su hospedador, E. coli. Este tipo de transducción recibió el nombre de transducción especializada, y sus caracteres distintivos son:sólo se transfieren marcadores cromosómicos cercanos al sitio de integración del ADN del fago (profago) en la célula lisogénica (p. ej., en el caso de l , los marcadores gal o bio);se produce únicamente como consecuencia de la inducción de la célula lisogénica por escisión del profago y consiguiente entrada a fase lítica, productora de nuevas partículas de fago;el ADN genómico de la bacteria transportado por la partícula transductora va unido a ADN del fago;la célula transductante se suele convertir en lisogénica para el fago correspondiente.





No hay comentarios:
Publicar un comentario