Funciona este grupo similar de túbulos y vesículas, localizado a mayor profundidad en el citoplasma cerca del aparato de Golgi, en la preparación de su contenido para la destrucción final de los lisosomas.
Los endosomas tempranos y tardíos constituyen el compartimiento endosómico. Las membranas de todos los endosomas contienen bombas de H ligadas al ATP que acidifican el interior de los endosomas al bombear de manera activa iones de H hacia ellos, de modo que el endosoma temprano tiene un PH de 6.0, en tanto que el tardío lo tiene de 5.5.
El material que entra en el endosoma temprano se puede recuperar desde dicho compartimiento y devolverse a su localización previa. Sucede así con los receptores de carga que necesitan recircular. Cuando la vesícula pinocítica se fusiona con el endosoma temprano, el ambiente ácido produce desacoplamiento del ligando desde su molécula receptora. El ligando se conserva dentro de la luz del endosoma temprano, en tanto que las moléculas receptoras (p. ej.: receptores de Lipo proteínas de baja densidad: LBD) se devuelven a la membrana plasmática. Algunos autores se refieren a este tipo de endosoma temprano como CDRL (Compartimiento para el desacoplamiento de receptor y ligando) (2-19e y 2-22t).
En plazo de pocos minutos después de entrar el endosoma temprano, el ligando se transfiere hacia un endosoma tardío (como en el caso de las lipoproteínas de baja densidad) o se empaca para devolverse hacia la membrana celular, sitio en el que descarga (p. ej.: transferrina) hacia el espacio extracelular. En ocasiones se transfieren tanto el receptor como el ligando (p. ej.: factor de crecimiento epidérmico y su receptor) hacia el endosoma tardío para su degradación final.
No ha podido dilucidarse el transporte entre los endosomas temprano y tardío. Algunos autores sugieren que los endosomas tempranos migran hacia una localización más profunda dentro de la célula y se vuelven endosomas tardíos. Otros sugieren que estos son dos compartimientos separados, y que hay vesículas endosómicas portadoras que transportan el material desde los endosomas tempranos hacia los tardíos. Se cree que se trata de grandes vesículas que contienen numerosas vesículas pequeñas, que se han identificado como cuerpos multivesiculares. Ambos conceptos reconocen la presencia de un sistema de microtúbulos a lo largo del cual el endosoma temprano o la vesícula endosómica portadora se abren camino hacia el endosoma tardío.
LISOSOMAS
El contenido de los lisosomas tardíos se descarga para la digestión enzimática en la luz de organitos especializados llamados lisosomas (2-22t y 2-23t). Cada lisosoma tiene forma redonda o es polimorfo. Si diámetro promedio es de 0.3 a 0.8 micras, y contiene por lo menos 40 tipos diferentes de hidrolasas ácidas, como sulfatasas, proteasas, nucleasas, lipasas y glucosidasas, etc. Como todas estas enzimas requieren un ambiente ácido para su funcionamiento óptimo, las membranas lisosómicas poseen bombas de protones que transportan de manera activa a los iones H hacia el lisosoma, con lo que conservan de esta manera su luz a un PH de 5.0 (fig. 2-19e).
Los lisosomas funcionan en la digestión no solo de macromoléculas, microorganismos fagocitados, desechos célulares y células, pues tambien lo hacen con los organitos en exceso o envejecidos como las mitocondrias y el RER.
Los lisosomas son orgánulos esféricos u ovalados que se localizan en el citoplasma celular.
En microscopía electrónica son fáciles de localizar porque es el orgánulo más oscuro (el más teñido) de cuantos contiene el citoplasma de la célula, mientras que las mitocondrias presentan una tinción más grisácea. La imagen de la izquierda muestra una célula completa, mientras que en la de la derecha se muestra una imagen parcial de la célula, los lisosomas se han señalado con flechas rojas.


El lisosoma consta de una membrana que contiene una cavidad o lumen, es un saco cerrado.
El contenido de los lisosomas en una sola célula es muy variable (fotos inferiores). Básicamente, el contenido de un lisosoma puede parecer homogéneo (como el de la primera imagen) o heterogéneo (como el resto).
| 1 | 2 | 3 | 4 | 5 | 6 |
 |  |  |  |  |  |
Las otras formas (los lisosomas secundarios,como el 2 y 3) son heterogéneos y pueden recibir nombres variados como lisosomas con formas mielínicas (4), cuerpos multivesiculares(5), o cuerpos residuales (6). Estos tipos se producen por efecto del almacenamiento en el lumen del lisosoma de sustancias que no pueden degradarse más o por la transformación en lisosomas de otro tipo de orgánulos como son los autofagosomas y los endosomas .
Los lisosomas son orgánulos donde se produce la degración de moléculas que provienen desde la endocitosis o del interior celular a partir de fagocitosis. Se diferencian de los endosomas porque no poseen receptores para la manosa 6 fosfato. Metchnikoff y sus colaboradores articularon a finales del siglo XIX la idea de que el material fagocitado era digerido en compartimentos intracelulares acidificados. Estos compartimentos fueron denominados lisosomas y aparecen en todas las células eucariotas. Son corpúsculos generalmente esféricos de dimensiones variables, con una unidad de membrana y pueden llegar a representar el 5 % del volumen celular, dependiendo de la tasa de digestión que se esté llevando en la célula.
El pH interno de los lisosomas es ácido, en torno a 5, y es en ese valor donde las enzimas lisosomales muestran su máxima actividad, por lo que se llaman hidrolasas ácidas. Se han encontrado aproximadamente 40 tipos de enzimas lisosomales que degradan proteínas (proteasas), lípidos (lipasas), sacáridos (glucosidasas) y nucleótidos (nucleasas). La membrana de los lisosomas protege al resto de la célula de esta actividad destructora, pero si ésta se rompiese el pH citoplasmático, próximo a 7,2, sería un obstáculo para la actividad de estas enzimas.
No todos los lisosomas son iguales y pueden contener juegos diferentes de enzimas. Cualquier defecto en alguna de las enzimas que existen en los lisosomas puede acarrear graves consecuencias, puesto que los productos que ellas deberían degradar quedarían almacenados en la célula como productos residuales. Por ejemplo, la enfermedad de la glucogenosis tipo II. En estos individuos la β-glucosidasa, que cataliza la degración del glucógeno, está ausente y por ello hay grandes acúmulos de glucógeno en los orgános, que suelen ser letales. Los lisosomas reciben distintos nombres según el estado de degradación de las moléculas que contienen: primarios, secundarios y cuerpos residuales. Los cuerpos residuales contienen material que ya no puede ser degradado y quedan almacenados en el interior celular o, como veremos más adelante, se fusionan con la membrana plasmática expulsando dicho material el medio extracelular.
Los lisosomas contienen transportadores de membrana específicos que van a permitir que los productos de la degradación, tales como aminoácidos, azúcares, nucleótidos, puedan ser transportados al citosol. También poseen en su membrana una bomba de protones para permitir su acidez interior. ¿Cómo se protegen las proteínas que se encuentran en la membrana de ser digeridas? Estas proteínas están fuertemente glucosidadas y parece que ello les proporciona protección.
Hay tres vías por donde llegan a los lisosomas las moléculas que se tienen que degradar:
a) Los lisosomas son considerados como la estación final de la vía endocítica. La mayoría de las moléculas que van a ser degradadas por esta vía tienen que pasar previamente por los endosomas. Las proteínas que no se reciclan de nuevo a la membrana plasmática o al TGN del aparato de Golgi desde los compartimentos endosomales son degradas en los lisosomas. La formación de los lisosomas es un asunto controvertido. Unos autores proponen que se forman por gemación o maduración a partir de los endosomas tardíos que ya contienen todas las enzimas degradativas necesarias así como las moléculas a degradar.Otros autores proponen que los lisosomas son orgánulos independientes de los endosomas y que reciben vesículas desde los endosomas o se producen fusiones entre endosomas tardíos y lisosomas.
Para que las proteínas integrales de la membrana plasmática sean dirigidas a los lisosomas se ha de producir una ubiquitinación de su parte citosólica, es decir, la adición de una molécula denominada ubiquitina. Ello es necesario para que las proteínas de membrana interaccionen con la maquinaria de reparto que se encuentran en los endosomas y no vuelvan a la membrana citoplasmática en vesículas de reciclado. Las interacciones con diversos complejos proteicos mantienen a las proteínas ubiquitinadas en zonas limitadas de la membrana endosomal, que poseen una cubierta en la que está presente la clatrina. Todo ello hace que sean retenidas en los endosomas tempranos y depués transportadas a los cuerpos multivesiculares, a los endosomas tardíos, y de ahí a los lisosomas, donde se degradan. Este mecanismo afecta a receptores, transportadores, canales, etcétera. Los receptores que no son ubiquitinados pero sí endocitados, cuando llegan a los endosomas tempranos suelen reciclarse hacia la membrana celular.
b) Las partículas obtenidas por fagocitosis siguen una vía propia. Las partículas como bacterias o restos celulares quedan en el interior celular englobadas por membrana formando un compartimento que madurará y se convertirá en el denominado fagosoma. La degradación de estas partículas se produce cuando se fusionan los fagosomas con los lisosomas.

Autofagia
c) Una tercera vía de llegada de moléculas a los lisosomas es la autofagia. Es un proceso ubicuo por el que los orgánulos deteriorados o material interno celular son eliminados.
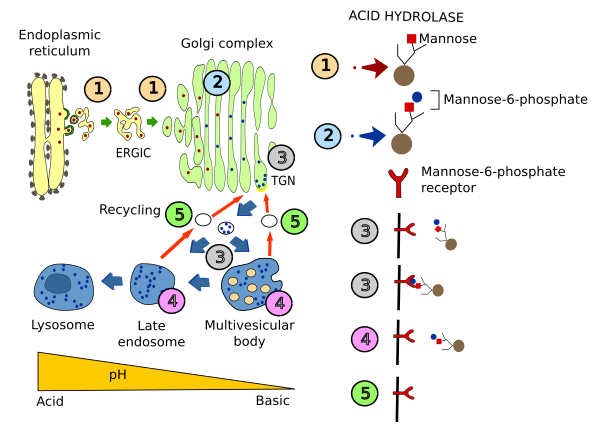
Pero además a los lisosomas han de llegar las hidrolasas ácidas encargadas de la degradación. Éstas se empaquetan en vesículas en el TGN del aparato de Golgi, las cuales se fusionarán con los endosomas tardíos y desde ahí llegan a los lisosomas. El mecanismo de selección de estas enzimas lo vimos en el apartado dedicado a los endosomas (ver figura =>). En el aparato de Golgi se añade a las enzimas lisosomales un grupo glucídico fosfatado, lamanosa-6-fosfato, que es reconocido por un receptor en el TGN del aparato de Golgi. La interacción del dominio citosólico de este receptor con la cubierta de clatrina permite englobar al receptor más la hidrolasa en vesículas que se dirigirán hacia los endosomas tardíos, y desde ahí hasta los lisosomas. Aunque éste sea el mecanimso principal existen otras proteínas que no requieren la fosforilación de las manosas para ir a los lisosomas. Estas proteínas son las integrales de la membrana. Estas proteínas contienen una secuencia de aminoácidos de destino que se encuentra en la cara citosólica de la proteína.
{kind=link}
Se ha creído tradicionalmente que los lisosomas tienen una intercomunicación muy limitada en la ruta vesicular cuando se compara con cualquier otro compartimento membranoso y se han considerado como un compartimento terminal. Durante los últimos años se han ido acumulando evidencias acerca de otra función de los lisosomas: su capacidad de participar en una exocitosis regulada. Por ejemplo, en el hígado se secretan enzimas lisosómicas a la bilis. También se ha observado la exocitosis de orgánulos con características similares a los lisosomas como es el caso de los melanocitos (los gránulos de melanina que pasarán a los queratinocitos que darán el color moreno a la piel). El acrosoma de los espermatozoides, una vesícula cargada de numerosas enzimas hidrolíticas, se libera durante la fecundación. Se ha propuesto desde hace tiempo que las células eucariotas son capaces de eliminar las sustancias que no pueden degradas más y esto sería posible si los lisosomas terminan por expulsar su material cuando se fusionan con la membrana plasmática. En las células de mamíferos donde sólo se produce secreción constitutiva se ha visto que bajo ciertas condiciones pueden realizar exocitosis regulada, por ejemplo, por una elevación de la concentración de calcio intracelular, lo cual ocurre, por ejemplo, durante las pequeñas roturas de la membrana citoplasmática, como vimos en el apartado dedicado a las membranas (Asimetría y reparación).
Orgánulos relacionados con los lisosomas (LRO lysosomal related organules)
Algunas células tienen orgánulos que se pueden relacionar con los lisosomas por su composición molecular y características fisiológicas, o son directamente lisosomas modificados. Entre ellos están los melanosomas de los melanocitos, gránulos azurófilos y basófilos de los leucocitos y mastocitos, gránulos líticos de los linfocitos T, gránulos densos de los megacariocitos, cuerpos lamelares de las células tipo II del pulmón, cuerpo Weibel-Palade de las células entoteliales y gránulos de los osteoclastos.
Los lisosomas reciben sus enzimas hidrolíticas lo mismo que sus membranas de la cara trans de Golgi; sin embargo, llegan en vesículas diferentes. Aunque ambos tipos de vesículas poseen una cubierta de clatrina que adquieren al desprenderse a manera de gemación desde la CTG, esta se pierde poco después de su formación. Las vesículas descubiertas se fusionan con los endosomas tardios.
Las vesículas portadoras de las enzimas lisosomales poseen receptores para la manosa-6-fosfato, a los cuales se fijan estas enzimas. En el ambiente ácido del endosoma tardío las enzima lisosomales se disocian de sus receptores y a continuación, su residuo de manosa se desfosforila, y los receptores recirculan para devolverse hacia la cara trans de Golgi. Debe entenderse que las hidrolasas lisosomales desfosforiladas no se pueden fijar ya a los receptores de manosa-6-fosfato y, por tanto, se quedan en el endosoma tardío (figs. 2-17t y 2-18e y 2-19e).
Cuando los endosomas tardíos poseen componentes tanto enzimáticos como de membrana, según algunos autores se fusionan con un lisosoma. Sin embargo, otros sugieren que maduran para convertirse en lisosomas.



No hay comentarios:
Publicar un comentario