Bacteriófagos
Un cianófago es un virus que infecta a las cianobacterias. El nombre se deriva a partir del término fago, para un virus cuyo blanco son las bacterias y cianobacterias para el tipo específico de bacteria que este virus utiliza para la replicación. Debido al importante papel de las cianobacterias como productores primarios en el mundo de los océanos, el estudio de su ecología fago es importante para el entendimiento global del ciclo del carbono.

Prochlorococcus myoviruses
Un Cianófago es un virus que infecta a las cianobacterias. El nombre se deriva a partir del término fago, para un virus cuyo blanco son las bacterias y cianobacterias para el tipo específico de bacteria que este virus utiliza para la replicación. Debido al importante papel de las cianobacterias como productores primarios en el mundo de los océanos, el estudio de su ecología fago es importante para el entendimiento global del ciclo del carbono.
Un Cianófago es un virus que infecta a las cianobacterias. El nombre se deriva a partir del término fago, para un virus cuyo blanco son las bacterias y cianobacterias para el tipo específico de bacteria que este virus utiliza para la replicación. Debido al importante papel de las cianobacterias como productores primarios en el mundo de los océanos, el estudio de su ecología fago es importante para el entendimiento global del ciclo del carbono.
Los virus no son únicamente patógenos, existen de todo tipo, hasta virus más chiquitos que infectan a virus más grandes. De hecho, la mayor parte de ellos infecta bacterias. Y si tenemos en cuenta que estas bacterias son responsables de la fijación y protagonistas en el ciclo de elementos clave como el carbono, el nitrógeno o el fósforo no sería arriesgado suponer que los virus también tienen un rol importante en labiósfera terrestre.
Hasta hace poco, los científicos creían que los virus eran súper específicos respecto a su huésped, pero ahora no están seguros. La evidencia más reciente sugiere que losvirus marinos no son tan exigentes. En 2003 se hicieron estudios que demostraron que un cierto tipo de cianófago no sólo infectaría distintas cianobacterias de un mismo tipo, sino cianobacterias en general. Entonces, los virus no estarían limitados a infectar a un solo genotipo, a una sola especie y tal vez ni siquiera a un solo género.
Teniendo en cuenta esto, los virus podrían tener un papel similar al de otros “pastores” del mar, como las protistas y el zooplancton, ya que las muertes selectivas de poblaciones microbianas estarían ligadas a una fluctuación dinámica entre virus y bacterias. Sin embargo, la evidencia de comportamientos aparejados entre virus y bacterias es limitada.
Cuando un virus mata a una bacteria, no sólo quedan los residuos de la célula huésped, sino que se liberan nutrientes orgánicos. Estos nutrientes pueden ser re absorbidos por otros microorganismos, así que la muerte de unos puede beneficiar indirectamente a otros. De hecho, antes de la muerte de estas células, los virus las modifican. Estas modificaciones ocurren a muchos niveles, también modificando el metabolismo, o sea que podrían estar “preparando” los nutrientes para que luego sean absorbidos con más facilidad por otros organismos. Esta teoría, llamada “primacía viral”, fue comprobada en modelos experimentales en aguas superficiales.
Dada la dificultad para cuantificar el rol que juegan los virus en ambientes complejos, los científicos se inclinan por estimarlo con modelos matemáticos. Para responder la pregunta: “¿Cómo se relaciona un grupo de bacterias con uno de virus?”, lo más importante es decidir dónde y cuándo los detalles importan y eso depende de qué quiera medir. Estos modelos desfragmentan las interacciones entre las bacterias y el ambiente, entre los virus y las bacterias y entre los virus y el ambiente.
Entonces, un modelo puede ignorar completamente la dinámica intracelular y concentrarse en los productos de estas interacciones, presentándolo como una reacción química, por ejemplo: 1 bacteria + 1 virus= 50 virus. Después, las poblaciones de virus y bacterias pueden derivarse de esta fórmula.
Un investigador de la Universidad de British Columbia estimó que los virus marinos podrían metabolizar 30 veces más carbono que el plancton. Esta capacidad de reciclar nutrientes sugiere que los virus deberían comenzar a ser incluidos en modelos cuantitativos y dinámicos de cambio climático. El ciclo del carbono en el mar aún no es comprendido del todo, por lo que la suma de pequeñas investigaciones podría ser muy importante a la hora de establecer un paradigma general.
La
conversión lisogénica es un proceso por el cual una
célula está lisogenizada, y expresa genes extra que el
bacteriofago porta, con lo cual se altera el
fenotipo de la célula huésped, otorgándole nuevas características.
La conversión lisogénica, o lisogenia, fue descubierta al inicio de 1950 cuando se observó que un grupo de cultivo de células bacterianas producía espontáneamente pequeñas cantidades de bacteriófagos.
En la conversión lisogénica, la introducción del genoma del fago en la célula bacteriana (estado lisógeno), le proporciona a la célula huésped una serie de nuevos caracteres que van desde cambios en la morfología colonial, hasta inhibición o intensificación de actividad enzimática, resistencia a antibióticos, inmunidad a la superinfección, así como cambios en las propiedades patogénicas, como la producción de toxinas.1
Cuando una célula lisogénica se divide, ambas células hijas son portadoras de profagos, heredando la lisogenia; y sólo un pequeño número de la población bacteriana se lisa espontáneamente liberándolos.
- En el caso de la cepa bacteriana inocua de Vibrio cholerae, producto de una conversión lisogénica, esta se transforma en una cepa tremendamente virulenta que puede causar cólera.
BACTERIÓFAGOSDr Gene Mayer Professor Emeritus
University of South Carolina School of Medicine
Traducido por :
Dr. en C. Paula Figueroa-Arredondo
|
|
|
|
|
|
|
|
|
|
OBJECTIVOS DE ENSEÑANZADescribir la composición general y la estructura del bacteriófago
Discutir el proceso infeccioso y la multiplicación en el cíclo lítico.
Explicar el cíclo lisogénico y su regulación.

© CellsAlive - James A. Sullivan |
INTRODUCTION
Definición
Los bacteriófagos (fagos) son parásitos intracelulares obligados que se multiplican al interior de las bacterias, haciendo uso de algunas o todas sus maquinarias biosintéticas (p. ej., los virus que infectan bacterias).
Existen muchas similaridades entre los bacteriófagos y los virus de células animales. Así, los bacteriófagos pueden ser visualizados como sistemas modelo de los virus de células animales. Además es necesario el conocimiento previo del ciclo de vida del bacteriófago para entender uno de los mecanismos por los cuales los genes bacterianos pueden transferirse de una bacteria a otra.
Alguna vez se pensó que el uso de los bacteriófagos podría ser una vía efectiva para tratar las infecciones bacterianas, pero pronto se hizo aparente que los fagos son removidos rápidamente del cuerpo así que resultaron de poco valor clínico. Sin embargo, los bacteriófagos son útiles en el diagnóstico de laboratorio para la identificación de bacterias patógenas (fago-tipificación). Aunque la fago-tipificación no se usa en el laboratorio clínico de rutina, sí se usa a en los laboratorios de referencia con propósitos epidemiológicos. Recientemente, se ha desarrollado un nuevo interés en el posible uso de los bacteriófagos para el tratamiento de infecciones bacterianas y en la profilaxis. De manera que la decisión de si los bacteriófagos serán usados en la medicina clínica o no, aún está por ser determinada.
|
PALABRAS CLAVEBacteriófago
Fago-tipificación
Cápside
Cola
Cortina contráctil
Placa de base
Fibras de la Cola
Fago virulento
Fase de eclipse
RNA mensajeros tempranos y tardíos
Placas
Pfu
Lisogenia
Fagos Temperados
Pro-fagos
Lisógenas
Extremos cohesivos
Recombination Sitio-específico
Represión
Inducción
Conversion Lisogénica
 Bacteriófago T4 (TEM x390,000) © Dennis Kunkel Microscopy, Inc. Used with permission Bacteriófago T4 (TEM x390,000) © Dennis Kunkel Microscopy, Inc. Used with permission
 Bacteriófago T4 Tinción Negativa micrografía electrónica © ICTV. Bacteriófago T4 Tinción Negativa micrografía electrónica © ICTV.
 Figura 1 Estructura del bacteriófago T4 Figura 1 Estructura del bacteriófago T4 |
COMPOSICIÓN Y ESTRUCTURA DEL BACTERIÓFAGO
Composición
Aunque diferentes bacteriófagos pueden contener diferentes materiales todos ellos contienen ácido nucleico y proteína.
Dependiendo del fago, el ácido nucléico puede ser ya DNA o ya RNA pero no ambos y puede existir en varias formas. Los ácidos nucléicos de los fagos a menudo contienen bases raras o modificadas. Estas bases modificadas protegen a los ácidos nucleicos del fago de las endonucleasas que cortan los ácidos nucléicos del huésped durante la infección. El tamaño de los ácidos nucleicos varía dependiendo del fago. Los fagos más simples solo tienen suficiente ácido nucleico para codificar un promedio de 3-5 productos génicos, mientras que los fagos mas complejos, pueden codificar para mas de 100 productos génicos.
El número de proteínas de diferentes clases y la cantidad de cada una de ellas en la partícula del fago variará dependiendo de la clase de fago que se trate. El fago mas simple posee varias copias de solo una o dos diferentes proteínas, mientras que los mas complejos podrían poseer muchos tipos de proteínas diferentes. La función de las proteínas durante la infección es proteger al ácido nucleico de las nucleasas de su medio ambiente.
Estructura
Los bacteriófagos vienen en muchas diferentes formas y tamaños. Las características básicas estructurales de los bacteriófagos se ilustran en la Figura 1, la cual muestra al fago denominado T4.
1. Tamaño - T4 está entre los fagos mas grandes, tiene aproximadamente 200 nm de largo y 80-100 nm de ancho. Otros fagos son mas pequeños. La mayoría de los fagos están entre un rango de 24-200 nm de longitud.
2. Cabeza o Cápside – Los fagos clásicos poseen una estructura a manera de cabeza y pueden variar de tamaño y forma. Algunos son icosaédricos (20 caras) otros son filamentosos. La cabeza o cápside está compuesta de muchas copias de una o más proteínas diferentes. Al interior de la cabeza se encuentra el ácido nucleico. La cabeza actúa como una cubierta protectora para el ácido nucleico.
3. Cola – Muchos, aunque no todos los fagos muestran una cola unida a la cabeza del fago. La cola es un tubo hueco a través del cual el ácido nucleico pasa durante la infección. El tamaño de la cola puede variar y algunos fagos ni siquiera la tienen. En los fagos más complejos como T4, la cola se rodea de una cortina contráctil durante la infección de la bacteria. Al extremo de la cola los fagos más complejos como T4 presentan una placa en la base y una o mas fibras unidas a ella. Esta placa de base y las fibras de la cola están involucradas en la unión de los fagos a la célula bacteriana. No todos los fagos tienen placas de base ni fibras de la cola, En tales casos existen otras estructuras que se ven asociadas en la unión de partícula del fago a la bacteria.
INFECCIÓN DE LAS CÉLULAS HUÉSPED
Adsorción
El primer paso en el proceso de infección es la adsorción del fago a la célula bacteriana- Este paso es reversible está mediado por las fibras de la cola o por alguna estructura análoga en aquellos fagos que carecen de las mismas. Las fibras de la cola se unen a receptores específicos en la célula bacteriana y la especificiad del huésped del fago (p. ej., la bacteria que es capaz de infectar) se determina usualmente por el tipo de fibras de la cola que un fago posee. La naturaleza del receptor bacteriano varía en las diferentes bacterias. Los ejemplos incluyen proteínas sobre la superficie externa de la bacteria, LPS, pili y lipoproteínas. Estos receptores están en la bacteria para otros propósitos y los fagos han evolucionado de manera que son capaces de usar estos receptores para llevar a cabo la infección.
Unión irreversible
La unión del fago con la bacteria vía las fibras de la cola es de naturaleza débil y es reversible. La unión irreversible del fago con la bacteria está mediada por uno o mas componentes de la placa de la base. Los fagos que carecen placas de la base tienen otras formas de unirse estrechamente a la célula bacteriana.
|
|
|
 Figura 2 Contracción de la cortina de la cola del fago T4 Figura 2 Contracción de la cortina de la cola del fago T4 |
Contracción de la cortina
La unión irreversible del fago a la bacteria da como resultado la contracción de la cortina (para aquellos fagos que la presentan) y la fibra hueca que es la cola se ve empujada a través de la envoltura bacteriana (Figure 2). Los fagos que no tienen cortinas contráctiles usan otros mecanismos para introducir la partícula del fago al interior de la envoltura de bacteriana. Algunos fagos tienen enzimas digetivas que degradan varios componentes de la envoltura bacteriana.
Inyección del Ácido Nucleico
Cuando el fago ha logrado atravesar la envoltura bacteriana el ácido nucleico que se encuentra en la cabeza pasa a través de la cola hueca y penetra la célula bacteriana. Usualmente el único componente del fago que realmente penetra la célula es el ácido nucleico. Los remanentes del fado permanecen en el exterior de la bacteria. Hay algunas excepciones para esta regla. Esto es diferente de los virus animales, en los cuales la mayoría de las partículas virales normalmente se introducen en la célula. Esta diferencia se debe probablemente a la incapacidad de la bacteria para envolver a la bacteria a fin de endocitar materiales.
|
 Figura 3 Cíclo de vida de un fago lítico Figura 3 Cíclo de vida de un fago lítico

 Figura 4 Ensayo de fago lítico Figura 4 Ensayo de fago lítico |
CICLO DE MULTIPLICACIÓN DEL FAGO
Fagos Líticos o Virulentos
Definición
Los fagos líticos o virulentos son fagos que solo pueden multiplicarse en bacterias y matan a la célula debiso a la lisis al término del ciclo de vida.
Cíclo de replicación viral
El ciclo de vida de un fago lítico se ilustra en la Figura 3.
a. Periodo de eclipse – Durante la fase de eclipse las partículas de fago no infeccioso se pueden encontrar ya sea en el interior o en el exterior de la célula bacteriana.
El ácido nucleico del fago se apodera de la maquinaria biosintéticas del huésped y se elaboran los mRNAs y las proteínas fago específicas. Existe una expresión ordenada de la síntesis macromolecular dirigida por el fago, tal como se observa en las infecciones por virus animales. Los mRNAs tempranos codifican para las proteínas tempranas, las cuales se necesitan para la síntesis del DNA de fagos y para apagar la biosíntesis del DNA, RNA y proteínas del huésped. En algunos casos las proteínas tempranas literalmente degradan el cromosoma del huésped. Luego de que el DNA del fago se sintetiza, entonces los mRNAs y las proteínas tardías se fabrican. Las proteínas tardías son las proteínas estructurales que conforman al fago, también son las proteínas necesarias para causar la lisis de la célula bacteriana.
b. Fase de acumulación intracelular – En esta fase el ácido nucléico y las proteínas estructurales de que están hechos, se ensamblan y las partículas infecciosas se acumulan dentro de la célula.
c. Lisis y Fase de liberación – Después de un tiempo la bacteria empieza a sufrir lisis y los fagos intracelulares se liberan al medio. El número de partículas liberadas por bacteria infectada puede ser tan alto como 1000.
Ensayos para determiner fagos líticos.
Ensayo en placa
Los fagos líticos se cuentan mediante ensayos de formación de placas. Una placa se define como el área clara que resulta de la lisis de bacterias (Figure 4). (Cada placa se origina de un solo fago infeccioso). A la partícula infecciosa que da origen a la placa se le llama una unidad formadora de placa (ufp o en inglés pfu).
|
 Figura 5. Circularización del cromosoma del fago: extremos cohesivos Figura 5. Circularización del cromosoma del fago: extremos cohesivos Figura 6 Recombination Sitio-específico Figura 6 Recombination Sitio-específico |
Fago lisogénico o Temperado
Definición
Los fagos lisogénicos o temperados son aquellos que bien pueden multiplicarse via el ciclo lítico o entran en un estado quiescente en la célula. En este estado quiescente la mayoría de los genes del fago no se transcriben; el genoma del fago existe en un estado reprimido. Al DNA del fago en este estado reprimido se le conoce como profago porque no es un fago pero posee el potencial para producir fagos. En la mayoría de los casos el DNA de fago realmente se integra en el cromosoma del huésped y se replica junto con el cromosoma del huésped y se transmite a las células hijas. La célula que alberga un profago no se ve negativamente afectada por la presencia del profago y el estado lisogénico puede persistir indefinidamente. A la célula que alberga un profago se le conoce comolisógena.
Eventos que conducen a la lisogenia
El fago prototipo: Lambda
a. Circularización del cromosoma del fago- El DNA de Lambda una molécula lineal de doble cadena con pequeñas regiones de cadena sencilla en los extremos 5’. Estas cadenas sencillas de los extremos son complementarias (extemos cohesivos) de manera que pueden aparearse y producir una molécula circular. En la célula, los extremos libres del círculo se pueden ligar para formar un círculo covalentemente cerrado como se ilustra en la Figura 5.
b. Recombinación Sitio-específico – Un evento de recombinación catalizado por una enzima codificada por el fago, ocurre entre un sitio particular en el DNA del fago circularizado y un sitio particular en el cromosoma del huésped. El resultado es la integración del DNA del fago en el cromosoma del huésped como se ilustra en la Figura 6.
c. Represion del genoma del fago – Una proteína codificada por el fago, la llamaremos el represor es una proteína que presenta afinidad y se une con un sitio particular en el DNA del fago que llamaremos el operador y apaga la transcripción de la mayoría de los genes del fago, EXCEPTO la del gene que dá lugar al represor.
El resultado es un genoma del fago pero reprimido estable el cual está integrado dentro del cromosoma de la célula huésped. Cada fago temperado solamente reprimirá la expresión de su propio DNA y no la de otro fago, así que la represión es muy específica (inmunidad a la superinfección con el mismo fago).
|
 Figura 7 Terminación de la lisogenia Figura 7 Terminación de la lisogenia Figura 8A Electro-micrografía de barrido (Scanning electron micrograph, SEM) de células de Escherichia coli con particulas de fago (las que aparecen como pequeños puntos blancos) unidas al exterior de las células. Figura 8A Electro-micrografía de barrido (Scanning electron micrograph, SEM) de células de Escherichia coli con particulas de fago (las que aparecen como pequeños puntos blancos) unidas al exterior de las células.
© Scott Kachlany, Cornell University Ithaca, New York, USA and The MicrobeLibrary
 Figura 8B Figura 8B
SEM de células de E. coli con envolturas celulares destruídas, debido presumiblemente a la liberación del fago. Luego de que los fagos se replican dentro de la células huésped estos deben ser liberados de las mismas. Esto ocurre a menudo debido mediante lisis de la célula.
© Scott Kachlany, Cornell University Ithaca, New York, USA and The MicrobeLibrary |
Eventos que conducen a la terminación de la Lisogenia
En cualquier momento en que la bacteria lisogénica quede expuesta a condiciones adversas, el estado lisogénico puede ser terminado. Este proceso se le llama inducción. Las condiciones que favorecen la terminación del estado lisogénico incluyen: desecación, exposición al la luz UV o radiación ionizante, exposición a químicos mutagénicos, etc. Las condiciones adversas que conducen a la producción de proteasas (proteína recA) la cual destruye a la proteína llamada represor. Esto a su vez conduce a la expresión de los genes del fago, al proceso inverso al proceso de integración, llamado escisión y al proceso de multiplicación lítica.
Ciclo Lítico vs Ciclo Lisogénico
La decisión de lambda de entrar en el ciclo lítico o en el ciclo lisogénico cuando este penetra en la célula inicialmente está determinada por la concentración dentro de la célula, tanto del represor, como de otra proteína del fago llamada cro. La proteína cro del fago, apaga la síntesis del represor y por lo tanto previene el establecimiento de la lisogenia. Las condiciones ambientales que favorecen la producción de cro conducirán al ciclo lítico mientras que aquellas que favorecen la producción del represor favorecerán la lisogenia.
Importancia de la of Lisogenia
a. Modelo de la transformación por virus animales – La lisogenia es un sistema modelo para la transformación que llevan a cabo los virus de células animales.
b. Conversión Lisogénica – Cuando la célula está lisogenizada ocasionalmente hay genes extra que el fago lleva y que se empiezan a expresar en la célula. Estos genes pueden cambiar las propiedades en la célula. Estos genes pueden cambiar las propiedades de la célula bacteriana. Este proceso se le llama conversión lisogénica o conversión por fago. Esto puede tener importancia clínica, por ej. Se ha demostrado que los fagos lisogénicos llevan genes que pueden modificar al antígeno O de Salmonella, el cual es uno de los antígenos principales ante los cuales está dirigida la respuesta inmune. La producción de toxina por Corynebacterium diphtheriae está mediada por un gen que es acarreado por un fago. Solo aquellas cepas que se han convertido por lisogenia son patógenos.
|

{kind=link}

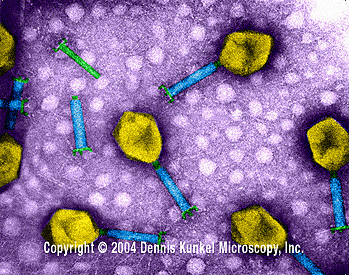 Bacteriófago T4 (TEM x390,000) © Dennis Kunkel Microscopy, Inc. Used with permission
Bacteriófago T4 (TEM x390,000) © Dennis Kunkel Microscopy, Inc. Used with permission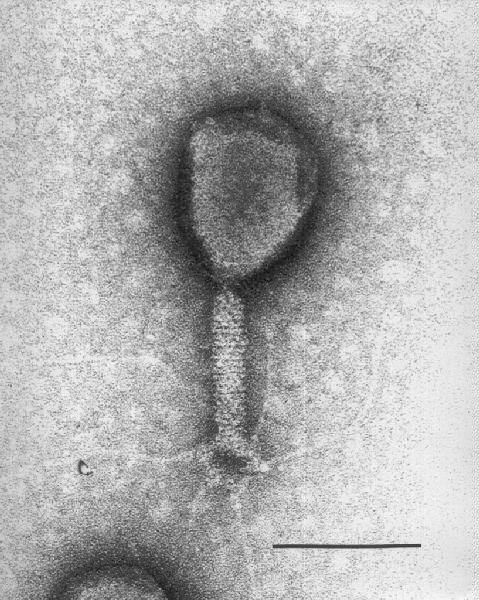 Bacteriófago T4 Tinción Negativa micrografía electrónica © ICTV.
Bacteriófago T4 Tinción Negativa micrografía electrónica © ICTV.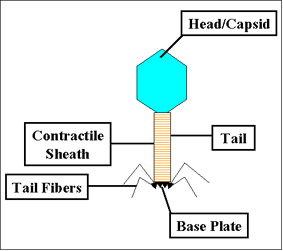 Figura 1 Estructura del bacteriófago T4
Figura 1 Estructura del bacteriófago T4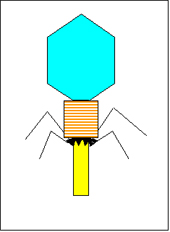 Figura 2 Contracción de la cortina de la cola del fago T4
Figura 2 Contracción de la cortina de la cola del fago T4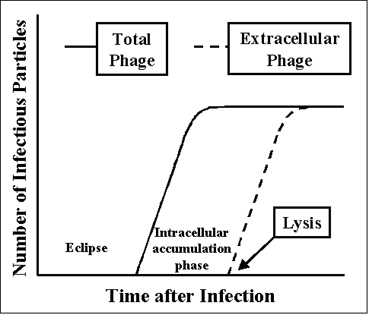 Figura 3 Cíclo de vida de un fago lítico
Figura 3 Cíclo de vida de un fago lítico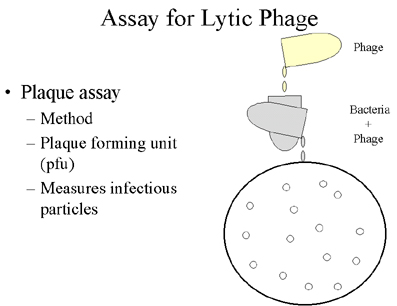 Figura 4 Ensayo de fago lítico
Figura 4 Ensayo de fago lítico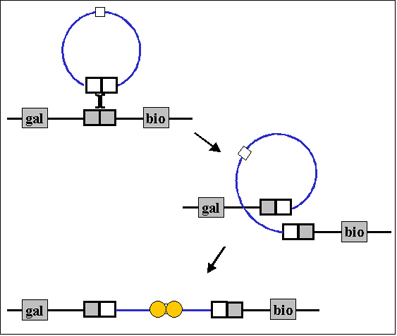 Figura 6 Recombination Sitio-específico
Figura 6 Recombination Sitio-específico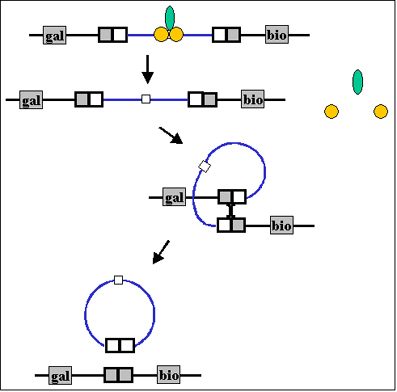 Figura 7 Terminación de la lisogenia
Figura 7 Terminación de la lisogenia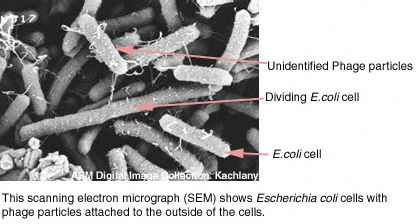 Figura 8A Electro-micrografía de barrido (Scanning electron micrograph, SEM) de células de Escherichia coli con particulas de fago (las que aparecen como pequeños puntos blancos) unidas al exterior de las células.
Figura 8A Electro-micrografía de barrido (Scanning electron micrograph, SEM) de células de Escherichia coli con particulas de fago (las que aparecen como pequeños puntos blancos) unidas al exterior de las células.
No hay comentarios:
Publicar un comentario