
Los discos imaginales son sacos de células epiteliales encontrados en las larvas de los insectos holometábolos. Representan los primordios de estructuras cuticulares del adulto que se formarán en la pupa durante la metamorfosis y que incluyen alas, patas, antenas, ojos, cabeza, tórax y genitales; están compuestos por grupos de células imaginales, determinadas pero indiferenciadas.12 Los discos imaginales han sido extensamente estudiados en Drosophila melanogaster.3
La mayoría de los órganos de la cabeza (ojos, antenas, cápsula cefálica, palpos) derivan de un solo tipo de disco. Estas estructuras se deben definir, dentro de un único conjunto de células (un solo disco), en lugares específicos, y su crecimiento y organización han de ser coordinados.
La producción de señales de comunicación intercelular y de factores de transcripción (los cuales regulan la expresión génica) en grupos celulares específicos y en momentos determinados del desarrollo es esencial para la diferenciación, crecimiento y organización de los tejidos. Está regulada por redes de genes que operan durante el desarrollo de los discos imaginales.
En Drosophila melanogaster se ha comprobado que los discos imaginales no se corresponden con metámerossino con porciones, anteriores o posteriores, de los metámeros. Estas porciones se denominan parasegmentos.4
Existen experimentos en los que se demuestra la especificidad y función de los discos imaginales. Tomando un disco imaginal de una larva donadora, por ejemplo de ojo, y trasplantándolo a una larva receptora en una región de la larva que va a dar lugar al abdomen, cuando la larva receptora del injerto termine su desarrollo y se convierta en imago, el adulto resultante tendrá un ojo extra en el abdomen.
Las células del disco imaginal están especificadas temprano en la embriogénesis, y el primordio está inicialmente compuesto de 20-50 células.5 El disco imaginal más grande, el del ala, contiene unas 60.000 células, mientras que los discos de la pata y el halterio contienen cerca de 10.000 células cada uno. Mientras que la mayoría de células de la larva tienen una capacidad mitótica muy limitada, los discos imaginales se dividen rápidamente en momentos específicos característicos. A medida que las células proliferan, forman un epitelio tubular que se pliega sobre sí mismo en una espiral compacta. Durante la metamorfosis, estas células proliferan, se diferencian y evierten.1
A medida que se desarrolla el disco, las células son restringidas a regiones de destino específico. Al final del tercer instar, justo antes de la pupación, el disco es un saco epitelial conectado por un delgado tallo a la epidermis de la larva; este tallo desempeña un papel importante en la posterior eversión del disco.5 Sobre un lado del saco el epitelio se enrosca en una serie de pliegues concéntricos. Durante la última etapa larval y la primera etapa pupal, el disco alcanza su tamaño final, y comienza a cambiar su morfología, y diferenciarse en su estructura adulta. A medida que comienza la pupación, las células del centro del disco se extienden hacia afuera para convertirse en las porciones más distales de la estructura, y las células externas se convierten en las estructuras proximales. Finalmente en la metamorfosis, el disco se somete a un proceso de eversión y se mueve del interior al exterior del animal.5
Después de diferenciarse, las células de los apéndices y de la epidermis secretan una cutícula apropiada para cada región específica. Aunque el disco está compuesto primariamente de células epidérmicas, un número pequeño de células adepiteliales migran hacia el disco en el desarrollo temprano. Durante el estadio pupal, estas células dan origen a los músculos y nervios de la estructura.1
La especificación de los destinos celulares generales se produce en el embrión. Los destinos celulares más específicos son especificados en los estadios larvales, a medida que las células proliferan.1
Formación de estructuras[editar]
Discos imaginales de la pata[editar]

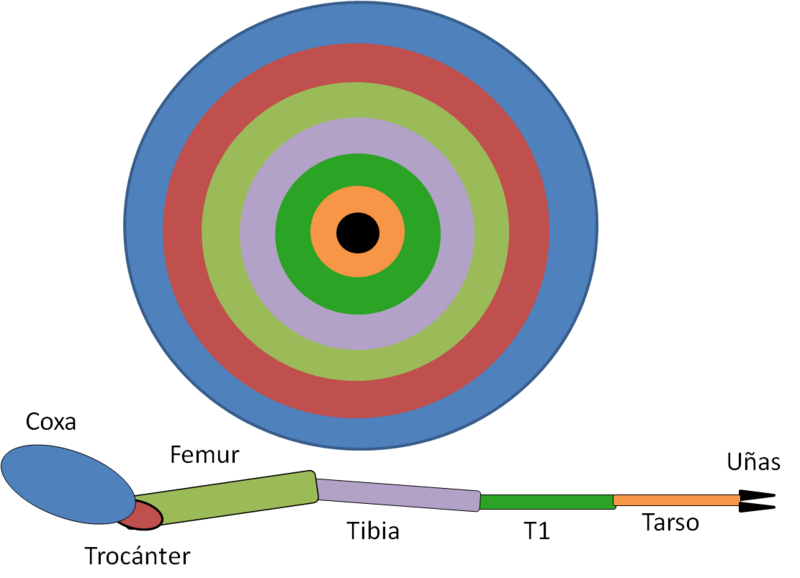
El tipo de estructura de la pata generada está determinado por las interacciones entre varios genes en el disco imaginal. El centro del disco secreta la concentración más alta de los morfógenos Wingless (Wg) y Decapentaplegic (Dpp), durante el tercer estadio larval del disco de la pata. Las concentraciones altas de los factores paracrinos provocan la expresión del gen Distal-less. Las concentraciones moderadas causan la expresión del gen dachshund y las concentraciones más bajas causan la expresión del gen homothorax. Aquellas células que expresan Distal-less se extienden hacia fuera para convertirse en las estructuras más distales de la pata – la garra y los segmentos tarsales; las que expresan homothorax se convierten en la estructura más proximal, la coxa. Las células que están expresando dachshund llegan a ser el fémur y la tibia proximal. Las áreas de superposición, entre homothorax y dachshund, producen el trocánter y la tibia distal. Estas regiones de expresión génica son estabilizadas por interacciones inhibitorias entre los productos proteicos de estos genes y de los genes vecinos.1
La diferenciación del disco requiere de una señal hormonal, una serie de pulsos de la hormona 20-hidroxiecdisona (20E). El primer pulso, administrado en los estadios larvales tardíos, da comienzo a la formación de la pupa, detiene la división celular en el disco y da comienzo a cambios en la forma celular que conducen la eversión de la pata, la cual se produce sin división celular y se debe principalmente a cambios en la forma celular dentro del epitelio del disco.1
Las estructuras de la pata se diferencian dentro de la pupa, de modo tal que en el momento en el que el insecto eclosiona, ellas están completamente formadas y funcionales.1
Discos imaginales de las alas[editar]
Los ejes del ala son especificados por los patrones de expresión de genes que dividen al embrión en compartimientos interactuantes diferenciados. En el primer estadio, la expresión del gen engrailed distingue el compartimiento posterior del ala del compartimiento anterior. El factor de transcripción Engrailed activa al gen hedgehog en el compartimiento posterior, Hedgehog actúa como un factor de transcripción de corto alcance para activar la expresión del gen decapentaplegic en un estrecho ámbito de células en la región anterior del disco del ala. Dpp actúa como un factor paracrino difusible de largo alcance, estableciendo un gradiente de concentración. Las altas concentraciones de Dpp activan a los genes spalt y oculomotor blind (omb) mientras que a bajas concentraciones solo se activa omb. Estos genes expresan los factores de transcripción que activan las proteínas que forman las regiones apropiadas del ala.1
El eje dorsoventral del ala se forma en el segundo estadio mediante la expresión del gen apterous en las células dorsales prospectivas del disco del ala. Aquí, la capa más superior del ala se distingue de la capa más inferior de la hoja del ala. El gen vestigial se mantiene “encendido” en la porción ventral del disco del ala. En el límite entre los compartimientos dorsal y ventral, los factores de transcripción Apterous y Vestigial interactúan para activan al gen wingless. Wingless actúa como un factor de crecimiento para promover la proliferación celular que extiende el ala, y también ayuda a establecer el eje proximal-distal del ala: niveles elevados de Wingless activan al gen Distal-less, que especifica las regiones más distales del ala.1
Discos imaginales del ojo-antena[editar]
Estos discos son particulares puesto que, a diferencia de los de las antenas y las patas, producen diferentes estructuras adultas. Estas regiones se subdividen en primordios que darán origen a antenas, ojos compuestos y regiones específicas de la cápsula cefálica. Aún no se ha entendido bien como se forman y diferencian estos primordios, ya que La mayoría de estudios se han enfocado en eventos tardíos en la diferenciación del disco.6
El surco morfogenético, que atraviesa el epitelio del disco comienza en el tercer instar larval y diferencia las células de la retina. Dpp, Wg y hh están involucrados en la regulación del progreso del surco. Experimentos con mutantes sugieren que la expresión de hh en la diferenciación de células fotorreceptoras induce la expresión de Dpp en células anteriores adyacentes y la progresión del surco. Aunque el rol del Dpp en la progresión del surco sea relativamente menos, su expresión temprana a lo largo de los márgenes posterior y lateral del disco es necesaria para la iniciación del surco. La formación del surco mediada por Dpp es antagonizada por Wg, que se expresa a lo largo de los márgenes dorsal y ventral del primordio del ojo.6
Además de su rol en la formación del surco, Dpp previene el destino de cabeza en el primordio de ojo. Esta función temprana de Dpp es mediada por la represión de Wg y el gen homeótico orthodenticle (odt). Mediante la represión de la expresión de Wg, Dpp permite el desarrollo del ojo compuesto. En ausencia de Dpp, el ojo es transformado en su destino por defecto, cutícula dorsal de la cabeza (frons).6
Además se requiere de hh en la progresión del surco morfogenético y en la diferenciación de los omatidios. hh juega un papel temprano en el patrón global del disco de ojo-antena. En ausencia de hh, Dpp no es expresado en el disco posterior de ojo. Como resultado, Wg y otd son expresados ectópicamente, causando una transformación de ojo a vértice.6
Transdeterminación[editar]
Artículo principal: Transdeterminación
Las células de un disco imaginal pueden experimentar transdeterminación, o cambiar su estado determinado al de otro tipo de disco.7
Muchos eventos de transdeterminación se asemejan a los efectos de mutaciones homeóticas. La transdeterminación de los discos imaginales puede ser inducida in situ por la no-expresión de proteínas homeodominio y la proteína nuclear Vestigial. Estas observaciones indican que la expresión localizada de estos factores reguladores es critica para mantener el destino celular determinado.7
Proliferación compensatoria[editar]
Cuando el daño a los discos imaginales es difuso, de tal manera que las células muertas están intercaladas con las células sobrevivientes, se incrementa la proliferación de las sobrevivientes. La proliferación compensatoria parece ocurrir por una combinación de proliferación acelerada así como una prolongación de la fase proliferativa del desarrollo del disco. La presencia de proliferación difusa, como ocurre durante el desarrollo normal, sugiere que la proliferación compensatoria puede estar regulada diferencialmente a regeneración basada en blastema pero puede estar gobernada por los mismos mecanismos de desarrollo que regula el crecimiento de tejido hasta lograr un disco de tamaño y forma específico.5
Los mecanismos por los que la proliferación compensatoria puede ser activada involucra que las células muertas emitan señales para estimular la proliferación de sus vecinos sobrevivientes.

El edeago o aedeagus es el órgano copulador intromitente de los insectos machos, a través del cual secretan el esperma desde los testículos durante la cópula con la hembra. El edeago es parte del abdomen del macho. Los testículos conectan con el edeago a través de los conductos deferentes. Forma parte del falo, una estructura compleja y a menudo con características diferenciales entre especies, compuesta de solapas y ganchos más o menos esclerotizados y que en algunas especies también incluye el pterigopodio, un par de órganos que facilitan el acoplamiento y sujeción del macho a la hembra durante la copulación. Durante la copulación, el edeago conecta con el oviporo de la hembra. El edeago puede ser bastante pronunciado o de tamaño mínimo.
La base del edeago puede ser la faloteca parcialmente esclerotizada, también denominada falosoma o teca.1 En algunas especies la faloteca contiene un espacio, llamado endosoma (bolsa de alojamiento interno), en su interior puede retraerse el extremo del edeago. Los conductos deferentes en ocasiones vierten en la faloteca junto a la vesícula seminal.23
El esperma de los artrópodos no pasa a la hembra en estado líquido con espermatozoides nadadores, sino que contiene cápsulas denominadas espermatóforos que encierran los espermatozoides. Además de los espermatóforos, en algunas especies el edeago también descarga spermatofilax, una bola de secreciones nutrientes para ayudar a la hembra a alimentar a la descendencia.
Los élitros son las alas anteriores, modificadas por endurecimiento (esclerotización), de ciertos órdenes de insectos como los coleópteros. Se denominan hemiélitros cuando el endurecimiento no es total sino parcial; su porción basal (próxima al pronoto) está engrosada y endurecida, y la porción apical o distal es membranosa (membrana del hemiélitro). Es típico del orden de los coleópteros.12

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Sirven como protección para las alas posteriores que están inmediatamente debajo y que sirven para volar. En algunos casos, los élitros están unidos, dejando al insecto sin capacidad de volar. Tienen asimismo una función de equilibrio durante el vuelo.
No hay comentarios:
Publicar un comentario