Lycophyta es una División de las plantas vasculares que se caracteriza por poseer una alternancia de generaciones bien manifiesta (con esporofito y gametofito de vida libre), con el esporofito formando un cormo (vástago, raíz y un sistema de haces vasculares que los vincula) de tipo primitivo: el vástago consta de tallo, y en las lycopsida también de hojas de tipo microfilo (de origen diferente de los megafilos de las euphyllophyta, probablemente derivadas de modificaciones de esporangios laterales, con un único haz vascular).
Las raíces adventicias son originadas del vástago, las raíces tienen una característica ramificación dicotómica. En el corte transversal del tallo y la raíz los haces vasculares forman un patrón ("estela") que al menos en las Lycopsida, en la raíz es una actinostela y en el tallo es derivada de la protostela con el xilema exarco, los esporangios son de inserción lateral, y en las lycopsida crecen en estrecha asociación con los microfilos estando ubicados en su cara adaxial, y usualmente con forma acorazonada.
Son las plantas vasculares más antiguas que existen en la actualidad, formando un clado (grupo monofilético) diferente de las euphyllophyta en donde se reúne el resto de las plantas vasculares. Si bien su diversificación parece haber comenzado hace 200 millones de años aproximadamente, la diversificación que generó las especies conocidas en la actualidad parece haber empezado recién hace unos 80 millones de años (al terminar el Cretácico). En la actualidad todavía existe la clase Lycopsida (el resto de las clases ya extintas), siendo sus géneros más sobresalientes Lycopodium, Selaginella e Isoetes.
Durante mucho tiempo se las ubicó junto con las Monilophyta o helechos, formando la división Pteridophyta, que hoy se sabe que es parafilética. Hasta hace poco tiempo se creía que las lycophytas eran tan antiguas como Psilotum y Equisetum, pero hoy se sabe que esos dos géneros son helechos que perdieron sus megafilos y otros caracteres de forma secundaria (después de haberlos adquirido).
Las Lycophytas fueron especialmente diversas y abundantes en el Carbonífero, y los restos de esas plantas representan nuestros mayores depósitos de carbón. Hoy en día hay unas 1.200 especies de lycophytas, que pertenecen a varios linajes diferentes.

El Clade Lycopsida
El clado Lycopsida, de los cuales los lycopsids de vida son miembros, también se separaron en el Devónico temprano. La planta Rhynie Chert, asteroxylon mackieies miembro de este clado. asteroxylon mackiei (Figura 5.3c) (VG 01:12) se caracteriza por una protostele lobulado, lateral, reniforme, acechado esporangios, y un eje cubierto de enaciones (Figura 5.3c) , colgajos multicelulares de tejido que emergen de la epidermis de la planta. En asteroxylon mackiei , enaciones no están vascularizados, sin embargo, una pequeña extensión del cilindro vascular se extiende a la base de cada enation. Es imposible decir cómo, o si, esta disposición podría haber funcionado. En su lugar, el cambio en la diferenciación de tejido vascular que condujo a la hebra vascular enation-Ward puede estar relacionada con el desarrollo de la propia enation, con un papel funcional para la vasculatura enation siguiente más tarde.
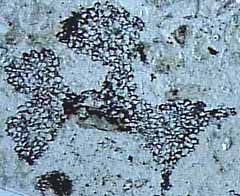{kind=link}
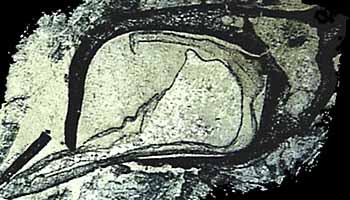
Sea cual sea su origen, la traza vascular en asteroxylon mackiei es una tentadora intermedio en la evolución de hojas verdaderas en el clado lycopsid. Verdaderos hojas del microfilos -son observados en baragwanathia y también en Drepanophycus (Figura 5.5) del Devónico temprano del hemisferio norte. Drepanophycus (VG 01:13) , como baragwanathia , tiene hojas vascularizados por una sola hebra no ramificado. Stalked, esporangios reniformes residen claramente en el eje de las hojas. Ejes puede ser de varios centímetros. de diámetro y rama irregular. Hojas de Drepanophycus surgen en un filotaxia más o menos espiral, pero el patrón de disposición de las hojas es variable y pueden aparecer casi verticiladas en algunos puntos a lo largo del eje. La variabilidad en la disposición apéndice es característico de los primeros miembros de varios subtipos diferentes y puede ser un indicio de la evolución de los mecanismos de regulación del desarrollo temprano en la historia de clados.
{kind=link}
 |
| Figura 5.5: Drepanophycus (A) la reconstrucción de toda la planta, (B) la sección transversal de estela lobulada. |
Junto con Drepanophycus , una variedad de otros lycopsids herbáceas floreció desde el Devónico Temprano en el Pennsylvania. Archaeosigillaria (VG 01:14) tiene ejes dicotomizar que eran de hasta 5 cm de diámetro. Microfilos están dispuestas en espiral y tienen lígulas , que une a estos grupos con vivir Isoetes y Selaginella , junto con los lycopsids árbol del Pennsylvanian. Archaeosigillaria tenido una actinostele exarca con escalariformes y reticulares traqueidas. Los esporangios nacieron adaxialmente en las hojas no modificados. Leclercqia tiene microfilos que ramificados en sus puntas (Figura 5.6, Figura 5.3D) (VG 01:15) , lo cual viola la definición técnica de micrófilo. Sin embargo, la cadena ramificada vascular se deriva claramente en este grupo. Leclercqia estela 's fue un actinostele exarca con escalariformes y pozos confinadas ovales en traqueidas metaxilema. Los esporangios nacen en tallos en la superficie adaxial de las hojas modificadas, otro personaje derivado en este linaje. Los esporangios se agrupan en zonas fértiles que alternan con las hojas vegetativas.
{kind=link}
{kind=link}
 | Figura 5.6: Leclercqia micrófilo. Un esporangio nació adaxialmente en cada hoja fértil; esporangio descansaba sobre un cojín de esporangios, proximal a la fosa lígula. |
Arborescente Lycopsids del Carbonífero
Debido a la variedad de estilos de conservación (por ejemplo, permineralizations bolas de carbón, compresiones / impresiones, yesos / moldes) en el que se registran, tenemos una excelente imagen de los lycopsids arborescentes que dominaron la turba a la droga y pantanos clásticas del Carbonífero. Estas plantas ampliamente estudiados tuvieron que ser minuciosamente reconstruido a partir de órganos dispersos. Debido a esto, podrás ver una gran cantidad de nombres genéricos de formulario asociados a cada organismo. En algunos casos, (sobre todo los tallos) forma única géneros parece corresponder a un solo género organismo. En otros casos (por ejemplo, follaje y microsporas), varios géneros madre pueden encontrarse en conexión orgánica con un solo género del órgano.
Algunos Ejemplos:Inicialmente, puede ser difícil "ver el árbol de los órganos" pero con el tiempo será capaz de reconstruir toda la planta.
- Lepidodendron - tallo (VG 2: 1)
- Lepidophloios - tallo (VG 2: 2)
- Sigillaria - vástago (VG 2: 3)
- Lepidophylloides - hojas (VG 2: 4) (VG 2: 5)
- Lepidostrobus - libre Sporing mono o conos bisporangiate (VG 2: 8) (VG 2: 9)
- Lepidocarpon - conos megaesporangiado (VG 2: 7)
- Stigmaria - rhizophores (VG 02:11) (VG 02:12)
- Lycospora - microsporas
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nota : La suerte quiso que la mayoría de los géneros madre tener prioridad nomenclatural (es decir, que fueron descritos por primera vez) por lo que puede ser usado para describir la planta entera cuando ha sido completamente reconstruido. Sin embargo, usted debe tener cuidado para entender si usted está hablando de Lepidodendron del género madre o Lepidodendron el árbol. ¿Por qué crees que sería importante mantener los nombres genéricos de forma aún cuando entendemos cómo debe ser reconstruido toda la planta? |
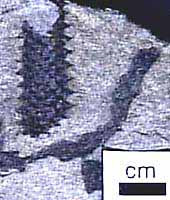
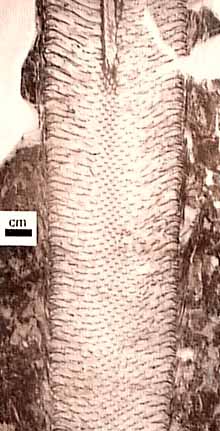
Árbol Lycopsids: El Lepidodendrales o lycopsids árbol (Figura 5.7) forman un clado de la diversidad Paleozoico tardío que la mayoría cree que es hermana de modernas Isoetes . Lycopsids arborescentes vivieron desde el Devónico tardío a través del Pérmico, con un cenit y una extinción importante en el Medio-Tardío de Pensilvania.
Lycopsids arborescentes comenzó la vida como un embrión en germinación de una megaspora indehiscente. Temprano en la vida, la fijación de las plántulas se caracteriza por una gruesa corteza que rodea a una esbelta prostele (Figura 5.7b). Este desarrollo probablemente dio la planta joven una apariencia muñón similares. Este patrón difiere de las plantas de semillas de árboles de tamaño, cuyo crecimiento primario es principalmente hacia arriba y la circunferencia expansión se produce con el tiempo por la acumulación de xilema secundario. Como el lycopsid arborescente creció más alto, la corteza adelgazada algo, se añadió una médula para formar un sifonostela, y el crecimiento secundario de un cambium vascular unifacial se inició. En algún momento en el desarrollo, el meristemo apical dicotomizada y continuó haciéndolo a intervalos. Sin embargo, más allá del punto de primera ramificación, ejes comenzaron a escasear, la médula disminuyó de diámetro, corteza adelgazada y el crecimiento termina en puntas de las ramas delgadas vascularizados por sólo un puñado de traqueidas dispuestos en una protostele. Comúnmente, los meristemos apicales diferencian en estructuras reproductivas, por lo que estas plantas monocárpica y determinante en su patrón de crecimiento.
   |
Figura 5.7: lycopsid arborescentes ( Lepidodendron ). (A) La ontogenia de un arborescente típica lycopsid; secciones (B) transversales de Lepidodendron tronco. ¿Cuál es la principal fuente de apoyo en este tipo de planta? |
Esta interpretación increíble fue desarrollado por DA Eggert (1961), quien reconstruyó minuciosamente el desarrollo de Lepidodendron seccionando en serie muchas personas preservadas en las bolas de carbón. A través de este trabajo detallado, reconoció que la base y la punta de lycopsids arborescentes mostrado una protostele, mientras que las porciones medias tenían siphonosteles claros con grandes médulas. Esta observación inicial condujo a una reconstrucción de la historia del desarrollo de la planta, lo que, a su vez, dio pistas sobre su historia de vida.


No hay comentarios:
Publicar un comentario