LISOSOMAS
| ||||||||
| « Endosomas | En células vegetales » | |||||||
|
Los lisosomas son orgánulos donde se produce degradación de moléculas.
La degradación es llevada a cabo por enzimas denominadas hidrolasas ácidas que tienen una alta actividad a pH ácido. Estos enzimas llegan a los lisosomas desde el TGN, con los endosomas tardíos siendo un paso intermedio. Hay tres vías para llegar a los endosomas: Endocítica: endosomas tempranos, cuerpos multivesiculares, endosomas tardíos y lisosomas Fagocitosis: fagosomas y fusión con los endosomas. Autofagia: orgánulos o contenido citosólico son englobados en vesículas o autofagosomas que se fusionan con los lisosomas. Los lisosomas pueden, bajo ciertas circustancias, liberar su contenido al exterior celular por exocitosis.
Las moléculas que se incorporan por endocitosis cuyo destino es la degradación serán conducidas hasta los lisosomas. Metchnikoff y sus colaboradores articularon a finales del siglo XIX la idea de que el material fagocitado era digerido en compartimentos intracelulares acidificados. Estos compartimentos fueron denominados lisosomas y aparecen en todas las células eucariotas. Son corpúsculos generalmente esféricos de dimensiones variables, con una unidad de membrana. Su pH interno es ácido, en torno a 5, y es en ese valor donde las enzimas lisosomales muestran su máxima actividad, por lo que se llaman hidrolasas ácidas. Se han encontrado aproximadamente 40 tipos de enzimas lisosomales que degradan proteínas (proteasas), lípidos (lipasas), sacáridos (glucosidasas) y nucleótidos (nucleasas). La membrana de los lisosomas protege al resto de la célula de esta actividad destructora, pero si ésta se rompiese el pH citoplasmático, próximo a 7,2, sería un obstáculo para la actividad de estas enzimas. No todos los lisosomas son iguales y pueden contener juegos diferentes de enzimas. Cualquier defecto en alguna de las enzimas que existen en los lisosomas puede acarrear graves consecuencias, puesto que los productos que ellas deberían degradar quedarían almacenados en la célula como productos residuales. Por ejemplo, la enfermedad de la glucogenosis tipo II. En estos individuos la β-glucosidasa, que cataliza la degración del glucógeno, está ausente y por ello hay grandes acúmulos de glucógeno en los orgános, que suelen ser letales. Los lisosomas reciben distintos nombres según el estado de degradación de las moléculas que contienen: primarios, secundarios y cuerpos residuales. Los cuerpos residuales contienen material que ya no puede ser degradado y quedan almacenados en el interior celular o, como veremos más adelante, se fusionan con la membrana plasmática expulsando dicho material el medio extracelular.
Los lisosomas contienen transportadores de membrana específicos que van a permitir que los productos de la degradación, tales como aminoácidos, azúcares, nucleótidos, puedan ser transportados al citosol. Tambiénposeen en su membrana una bomba de protones para permitir su acidez interior. ¿Cómo se protegen las proteínas que se encuentran en la membrana de ser digeridas? Estas proteínas están fuertemente glucosidadas y parece que ello les proporciona protección.
Hay tres vías por donde llegan a los lisosomas las moléculas que se tienen que degradar:
a) Los lisosomas son considerados como la estación final de la vía endocítica. La mayoría de las moléculas que van a ser degradadas por esta vía tienen que pasar previamente por los endosomas. Las proteínas que no se reciclan de nuevo a la membrana plasmática o al TGN del aparato de Golgi desde los compartimentos endosomales son degradas en los lisosomas. La formación de los lisosomas es un asunto controvertido. Unos autores proponen que se forman por gemación o maduración a partir de los endosomas tardíos que ya contienen todas las enzimas degradativas necesarias así como las moléculas a degradar.Otros autores proponen que los lisosomas son orgánulos independientes de los endosomas y que reciben vesículas desde los endosomas o se producen fusiones entre endosomas tardíos y lisosomas.
Para que las proteínas integrales de la membrana plasmática sean dirigidas a los lisosomas se ha de producir una ubiquitinación de su parte citosólica, es decir, la adición de una molécula denominada ubiquitina. Ello es necesario para que las proteínas de membrana interaccionen con la maquinaria de reparto que se encuentran en los endosomas y no vuelvan a la membrana citoplasmática en vesículas de reciclado. Las interacciones con diversos complejos proteicos mantienen a las proteínas ubiquitinadas en zonas limitadas de la membrana endosomal, que poseen una cubierta en la que está presente la clatrina. Todo ello hace que sean retenidas en los endosomas tempranos y depués transportadas a los cuerpos multivesiculares, a los endosomas tardíos, y de ahí a los lisosomas, donde se degradan. Este mecanismo afecta a receptores, transportadores, canales, etcétera. Los receptores que no son ubiquitinados pero sí endocitados, cuando llegan a los endosomas tempranos suelen reciclarse hacia la membrana celular.
b) Las partículas obtenidas por fagocitosis siguen una vía propia. Las partículas como bacterias o restos celulares quedan en el interior celular englobadas por membrana formando un compartimento que madurará y se convertirá en el denominado fagosoma. La degradación de estas partículas se produce cuando se fusionan los fagosomas con los lisosomas.
 Autofagia
c) Una tercera vía de llegada de moléculas a los lisosomas es la autofagia. Es un proceso ubicuo por el que los orgánulos deteriorados o material interno celular son eliminados.
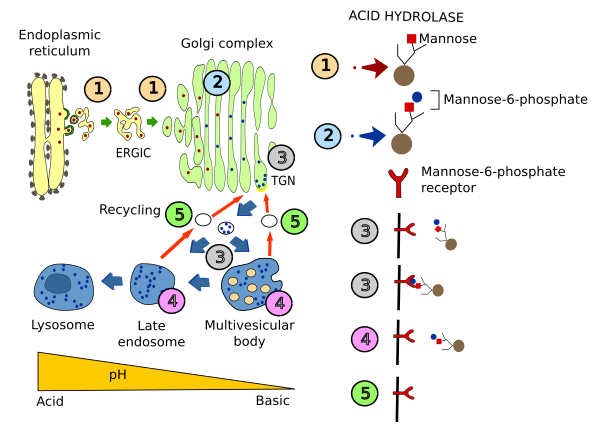
Pero además a los lisosomas han de llegar las hidrolasas ácidas encargadas de la degradación. Éstas se empaquetan en vesículas en el TGN del aparato de Golgi, las cuales se fusionarán con los endosomas tardíos y desde ahí llegan a los lisosomas. El mecanismo de selección de estas enzimas lo vimos en el apartado dedicado a los endosomas (ver figura =>). En el aparato de Golgi se añade a las enzimas lisosomales un grupo glucídico fosfatado, la manosa-6-fosfato, que es reconocido por un receptor en el TGN del aparato de Golgi. La interacción del dominio citosólico de este receptor con la cubierta de clatrina permite englobar al receptor más la hidrolasa en vesículas que se dirigirán hacia los endosomas tardíos, y desde ahí hasta los lisosomas. Aunque éste sea el mecanimso principal existen otras proteínas que no requieren la fosforilación de las manosas para ir a los lisosomas. Estas proteínas son las integrales de la membrana. Estas proteínas contienen una secuencia de aminoácidos de destino que se encuentra en la cara citosólica de la proteína.
Se ha creído tradicionalmente que los lisosomas tienen una intercomunicación muy limitada en la ruta vesicular cuando se compara con cualquier otro compartimento membranoso y se han considerado como un compartimento terminal. Durante los últimos años se han ido acumulando evidencias acerca de otra función de los lisosomas: su capacidad de participar en una exocitosis regulada. Por ejemplo, en el hígado se secretan enzimas lisosómicas a la bilis. También se ha observado la exocitosis de orgánulos con características similares a los lisosomas como es el caso de los melanocitos (los gránulos de melanina que pasarán a los queratinocitos que darán el color moreno a la piel). El acrosoma de los espermatozoides, una vesícula cargada de numerosas enzimas hidrolíticas, se libera durante la fecundación. Se ha propuesto desde hace tiempo que las células eucariotas son capaces de eliminar las sustancias que no pueden degradas más y esto sería posible si los lisosomas terminan por expulsar su material cuando se fusionan con la membrana plasmática. En las células de mamíferos donde sólo se produce secreción constitutiva se ha visto que bajo ciertas condiciones pueden realizar exocitosis regulada, por ejemplo, por una elevación de la concentración de calcio intracelular, lo cual ocurre, por ejemplo, durante las pequeñas roturas de la membrana citoplasmática, como vimos en el apartado dedicado a las membranas (Asimetría y reparación).
| |||||||
domingo, 26 de abril de 2015
Atlas de histología vegetal y animal


{kind=link}
Suscribirse a:
Enviar comentarios (Atom)
No hay comentarios:
Publicar un comentario