- Neutralización.- La unión con el anticuerpo inhabilita al antígeno para ejercer el efecto tóxico que pudiera tener. Algunas toxinas bacterianas no pueden unirse a sus receptores en la superficie celular cuando están unidas a su anticuerpo específico. Muchos virus, aunque son complejos supramoleculares también pueden resultar neutralizados el mismo modo por anticuerpos específicos
- Precipitación.- La formación de grandes entramados antígeno-anticuerpo impide a ambos tipos de moléculas permanecer en disolución, dando lugar a un precipitado.
En cualquier caso el destino final de los complejos antígeno-anticuerpo es ser fagocitados y digeridos por las células fagocitarias (micrófagos y macrófagos). Así, el mecanismo de defensa específico de la respuesta inmunizante humoral, interactúa con el mecanismo de defensa inespecífico de las células fagocitarias, que complementa la acción del primero. La porción carboxi-terminal de las cadenas pesadas de las inmunoglobulinas (fragmento Fc), que no está implicada en la formación de los complejos antígeno-anticuerpo, es reconocida por receptores de membrana de las células fagocitarias. Ello permite a estas células fagocitar los complejos antígeno-anticuerpo mediante endocitosis mediada por receptores específicos que es mucho más eficaz que la endocitosis convencional.
Cuando los antígenos se encuentran anclados a la superficie de microorganismos invasores la formación de complejos antígeno-anticuerpo sobre dicha superficie tiene las siguientes consecuencias:
- Activación del sistema del complemento.- El conjunto de proteínas que integra el sistema del complemento, que también participa en los mecanismos de defensa inespecíficos, provoca lisis celular de los microorganismos invasores reconocidos por los anticuerpos.
- Opsonización.- El microorganismo cuyos antígenos de superficie han sido reconocidos por los anticuerpos queda literalmente erizado de fragmentos Fc de las inmunoglobulinas Estos fragmentos, como hemos comentado con anterioridad, constituyen señales que las células fagocitarias reconocen y utilizan para mejorar su eficacia. También en este caso los mecanismos específicos e inespecíficos cooperan en la defensa del organismo.
4.1.3.- ORIGEN DE LA DIVERSIDAD DE LOS ANTICUERPOS.
Uno de los aspectos más intrigantes del fenómeno de la respuesta inmunitaria fue durante mucho tiempo el origen de la gran diversidad de anticuerpos necesaria para poder reconocer a un número potencialmente infinito de antígenos. Puesto que los anticuerpos son proteínas, la explicación más acorde con la ortodoxia de la teoría “un gen – un polipéptido” apuntaba a que debía existir un número elevadísimo de genes codificadores de inmunoglobulinas, tanto para las cadenas pesadas como para las ligeras. De acuerdo con esta explicación, conocida a mediados del siglo XX como teoría germinal, una parte muy significativa del genoma estaría dedicada a la codificación de las inmunoglobulinas (los cálculos apuntaban a una cifra en torno a los 100.000 genes en la especie humana, cuando hoy sabemos que el total de genes no supera los 30.000). Una explicación alternativa, conocida como teoría de la variación somática apuntaba a un número de genes mucho menor, que mediante mecanismos de mutación y/o recombinación generaría durante la maduración de los linfocitos el amplio repertorio de anticuerpos requerido. Un inconveniente de esta teoría consistía en que no explicaba satisfactoriamente la presencia de regiones variables y regiones constantes en las inmunoglobulinas.
En 1965 W.J. Dreyer y J.C. Bennett propusieron un modelo para explicar la diversidad de los anticuerpos según el cual las regiones variables y las regiones constantes, tanto de las cadenas pesadas como de las ligeras, estaban codificadas por genes diferentes. Estos genes se encontrarían separados en las células de la línea germinal, pero durante la maduración de los linfocitos B el genoma sufriría distintas reordenaciones que colocarían adyacentes los fragmentos codificantes para uno y otro tipo de región, dando lugar a un mensaje continuo que codificaría una cadena completa. Además, existiría, de acuerdo con la teoría germinal, un elevado número de genes
 Las ideas de Dreyer y Bennett fueron consideradas “heréticas” en su momento pues contradecían algunos dogmas de la naciente biología molecular. Sin embargo, cuando diez años más tarde Susumu Tonegawa (Figura 21.9) dispuso de las herramientas adecuadas para someterlas a la experimentación, se comprobó que no sólo apuntaban en la dirección correcta, sino que sus autores se habían quedado cortos en su apreciación de un mecanismo que resultó ser mucho más complejo y sofisticado.
Las ideas de Dreyer y Bennett fueron consideradas “heréticas” en su momento pues contradecían algunos dogmas de la naciente biología molecular. Sin embargo, cuando diez años más tarde Susumu Tonegawa (Figura 21.9) dispuso de las herramientas adecuadas para someterlas a la experimentación, se comprobó que no sólo apuntaban en la dirección correcta, sino que sus autores se habían quedado cortos en su apreciación de un mecanismo que resultó ser mucho más complejo y sofisticado.
Tonegawa demostró que la información genética que codifica la secuencia de aminoácidos de las inmunoglobulinas se encuentra en las células germinales fragmentada en una serie de segmentos génicos. Estos segmentos constituyen familias multigénicas encargadas de codificar los distintos tipos de cadenas (Figura 21.10). Hay tres familias principales, que se encuentran situadas en diferentes cromosomas: una que codifica las cadenas ligeras κ, otra las cadenas ligeras λ y una tercera las cadenas pesadas. Cada familia se compone de varios tipos de segmentos (V, D, J, C y L), cada uno de los cuales puede aparecer en diferentes versiones o subtipos (p. ej. V1, V2, V3, etc.). Los segmentos de las familias V, D y J codifican las regiones variables; los segmentos C codifican las regiones constantes y los segmentos L codifican un péptido señal presente en las cadenas recién sintetizadas que se elimina posteriormente.
 Durante el proceso de maduración de los linfocitos B los segmentos codificantes sufren reordenaciones, diferentes en cada línea individual, de manera que la secuencia de DNA que finalmente codificará una cadena de una inmunoglobulina se ensambla tomando segmentos de las distintas familias siguiendo una pauta característica de cada tipo de cadena (H o L). Así, las cadenas ligeras muestran una secuencia de segmentos Vi+Ji+Ci, mientras que las pesadas muestran una secuencia Vi+Ji+Di+Ci, donde el subíndice indica la variante incluida en cada tipo de segmento. De este modo, con un número discreto de segmentos diferentes (en realidad unos pocos centenares) se puede formar un número elevado de combinaciones que presentarían especificidades para antígenos diferentes. Los segmentos del tipo C presentan pocas variantes y no afectan a la especificidad del anticuerpo, sino que son las que determinan el isotipo al que pertenece.
Durante el proceso de maduración de los linfocitos B los segmentos codificantes sufren reordenaciones, diferentes en cada línea individual, de manera que la secuencia de DNA que finalmente codificará una cadena de una inmunoglobulina se ensambla tomando segmentos de las distintas familias siguiendo una pauta característica de cada tipo de cadena (H o L). Así, las cadenas ligeras muestran una secuencia de segmentos Vi+Ji+Ci, mientras que las pesadas muestran una secuencia Vi+Ji+Di+Ci, donde el subíndice indica la variante incluida en cada tipo de segmento. De este modo, con un número discreto de segmentos diferentes (en realidad unos pocos centenares) se puede formar un número elevado de combinaciones que presentarían especificidades para antígenos diferentes. Los segmentos del tipo C presentan pocas variantes y no afectan a la especificidad del anticuerpo, sino que son las que determinan el isotipo al que pertenece.
Las reordenaciones se llevan a cabo por la acción de un equipo de enzimas específicos que cortan y empalman los segmentos de DNA. Los lugares de corte están señalizados por unas secuencias específicas denominadas secuencias señalizadoras de recombinación (RSS).
Adicionalmente, se ha detectado en linfocitos B con las secuencias codificantes para inmunoglobulinas ya reordenadas un fenómeno conocido como hipermutación somática, que ocurre tras el primer contacto del antígeno con los receptores de membrana del linfocito. El fenómeno consiste en un aumento espectacular de la tasa de mutación en las secuencias que codifican las regiones variables de las inmunoglobulinas. Las mutaciones introducen ligeras variaciones en la conformación de las regiones variables que repercuten en su afinidad por el antígeno. Posteriormente se seleccionan para generar la respuesta inmunitaria aquellas líneas de linfocitos B portadoras de las mutaciones que confieren una mayor afinidad.
4.2.- RESPUESTA INMUNITARIA CELULAR.
La respuesta inmunitaria humoral por sí sola constituye una poderosa barrera defensiva contra los microorganismos patógenos siempre que éstos se encuentren libres en el medio interno del organismo receptor. Sin embargo se muestra poco eficaz frente a parásitos intracelulares como los virus y algunas bacterias y protozoos, a las que los anticuerpos circulantes no tienen acceso, al menos durante alguna fase de su ciclo vital. Para solventar ese problema algunos vertebrados se han dotado de un sistema que funciona en paralelo con la inmunidad humoral, con la que además interactúa. Es la respuesta inmunitaria celular.
La respuesta inmunitaria celular es un complejo mecanismo diseñado para localizar y destruir células del propio organismo receptor que hayan sido invadidas por parásitos intracelulares. También puede actuar sobre células cancerosas y sobre tejidos ajenos al organismo receptor como ocurre en el caso de los transplantes de órganos.
Las células responsables de la respuesta inmunitaria celular son un tipo de leucocitos que tienen su origen en la médula ósea pero que maduran en un órgano linfoide llamado timo. En referencia a este órgano se denominan linfocitos T. En el ser humano el timo entra en regresión a partir de la adolescencia y desde entonces los linfocitos T maduran en otros órganos linfoides diseminados. La respuesta generada por los linfocitos T consiste en la liberación de una serie de sustancias citotóxicas, que destruyen células infectadas, o de mensajeros intercelulares, llamados citoquinas, que inducen la proliferación y estimulan la actividad e otras células del sistema inmunitario.
 Los linfocitos T presentan en la cara externa de sus membranas unos complejos receptores de las células T (TCR) análogos a los correspondientes de los linfocitos B (Figura 21.11). El componente principal de los TCR es una proteína de estructura similar a la de las inmunoglobulinas, formada por dos cadenas polipeptídicas cada una de las cuales tiene una región constante y una región variable. Esta proteína del TCR, al igual que las inmunoglobulinas de membrana de los linfocitos B, es la responsable de reconocer al antígeno en la etapa inicial de la respuesta inmunitaria celular.
Los linfocitos T presentan en la cara externa de sus membranas unos complejos receptores de las células T (TCR) análogos a los correspondientes de los linfocitos B (Figura 21.11). El componente principal de los TCR es una proteína de estructura similar a la de las inmunoglobulinas, formada por dos cadenas polipeptídicas cada una de las cuales tiene una región constante y una región variable. Esta proteína del TCR, al igual que las inmunoglobulinas de membrana de los linfocitos B, es la responsable de reconocer al antígeno en la etapa inicial de la respuesta inmunitaria celular.
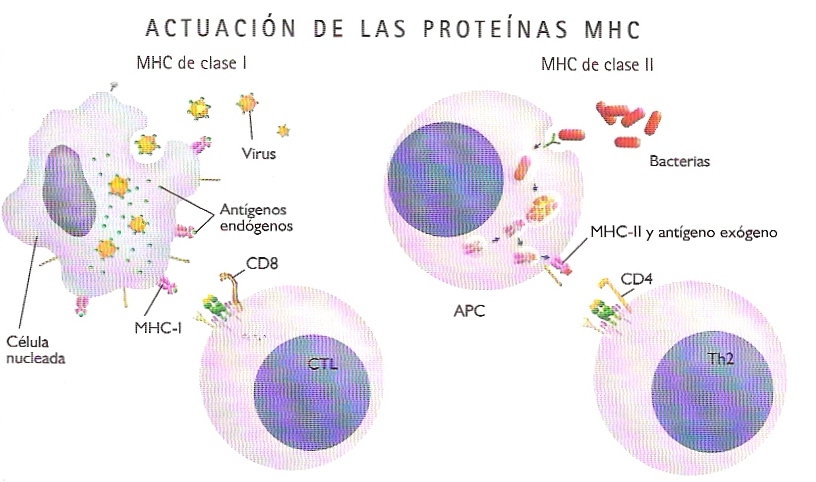
El proceso de reconocimiento del antígeno es en la respuesta inmunitaria celular mucho más complejo que en la humoral. En él interviene un conjunto de proteínas del organismo receptor y los genes que las codifican que se conocen con el nombre global de complejo principal de histocompatibilidad (MHC). Las proteínas MHC forman parte de la membrana celular y su misión es exponer fragmentos de antígenos sobre la superficie de la célula, de manera que éstos puedan ser reconocidos por los TCR de los linfocitos T (Figura 21.11). Los TCR, a diferencia de los BCR, no pueden reconocer antígenos libres, sino fragmentos de éstos (péptidos cortos) que se encuentran unidos a proteínas MHC (hecho que se conoce como restricción por el MHC).
Hay dos tipos de proteínas MHC:
- Proteínas MHC de clase I.- Se encuentran en todas las células nucleadas del organismo receptor. Reconocen y exponen sobre la superficie celular fragmentos antigénicos de origen endógeno, es decir sintetizados dentro de las propias células. Estos fragmentos pueden ser de proteínas víricas, si la célula se encuentra infectada por un virus, o proteínas tumorales de una célula cancerosa.
- Proteínas MHC de clase II.- Se encuentran exclusivamente en las llamadas células presentadoras de antígeno (APC) del sistema inmunitario. Las principales células presentadoras de antígeno son los macrófagos, que fagocitan de manera inespecífica microorganismos invasores, y los propios linfocitos B, que también pueden fagocitar los antígenos que desencadenan en ellos la respuesta humoral.
Existen dos tipos principales de linfocitos T, responsables de sendas modalidades de respuesta inmunitaria celular (Figura 21.10):
{kind=link}
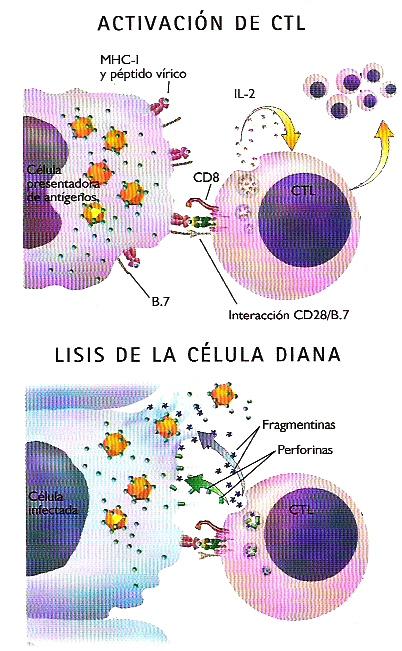
- Linfocitos T citotóxicos.- Reconocen mediante sus TCR fragmentos antigénicos presentados por proteínas MHC de clase I ancladas en la superficie de células infectadas por virus o que se han malignizado. El reconocimiento del antígeno desencadena en el linfocito T un doble respuesta: por una parte, el linfocito secreta unas citoquinas, denominadasinterleucinas, que inducen la proliferación de linfocitos T con la misma especificidad antigénica; por otra, el linfocito mata a la célula infectada inyectándole enzimas citotóxicas como las fragmentinas, que desencadenan en la célula infectada los mecanismos de muerte celular programada (apoptosis), y las perforinas, que abren poros en la membrana plasmática induciendo la lisis celular por fenómenos osmóticos.
- Linfocitos T auxiliares.- Reconocen mediante sus TCR fragmentos antigénicos presentados por proteínas MHC de clase II ancladas en la superficie de células presentadoras de antígenos. La interacción entre ambos tipos celulares desencadena en los linfocitos T auxiliares una respuesta consistente en la liberación de distintos tipos de citoquinas, que estimulan la proliferación de los propios linfocitos así como la actividad de las células presentadoras. En unos casos los fragmentos antigénicos son presentados por un macrófago y proceden de la digestión parcial de microorganismos invasores fagocitados por éste. La respuesta generada estimula por una parte la proliferación del clon de linfocitos T auxiliares implicado, y por otra aumenta la capacidad de los macrófagos para destruir por completo al germen estimulando la producción de lisosomas. En otros casos los fragmentos antigénicos son presentados por un linfocito B y proceden de la digestión de antígenos libres que ya han generado una respuesta de tipo humoral. La respuesta en esta ocasión consiste en la liberación de interleucinas por el linfocito T auxiliar, que inducen la proliferación del clon de linfocitos B específico del antígeno, aumentando así la producción de anticuerpos circulantes.
{kind=link}
5.- INTEGRACIÓN DE LOS MECANISMOS DE DEFENSA.
La discusión precedente demuestra que los distintos tipos mecanismos defensivos que han desarrollado los animales frente a los agentes infecciosos no funcionan aislada e independientemente unos de otros, sino más bien como partes complementarias e interactuantes de un todo integrado. Es posible que las distinciones entre mecanismos específicos e inespecíficos, o entre inmunidad humoral e inmunidad celular, sean más artefactos académicos que otra cosa y que, aunque se les pueda reconocer un cierto valor a la hora de sistematizar el conocimiento inmunológico, no estén reflejando adecuadamente una realidad mucho más compleja de lo que esas distinciones dibujan.
6.- MEMORIA INMUNITARIA.
Una de las propiedades más relevantes del sistema inmunitario es su capacidad para registrar los antígenos que han desencadenado una respuesta por su parte de tal modo que si posteriormente se produce un nuevo contacto con uno de ellos la respuesta generada es de mayor amplitud y eficacia. El mecanismo que explica esta propiedad se inscribe en el marco de lateoría de la selección clonal, que analizaremos a continuación.
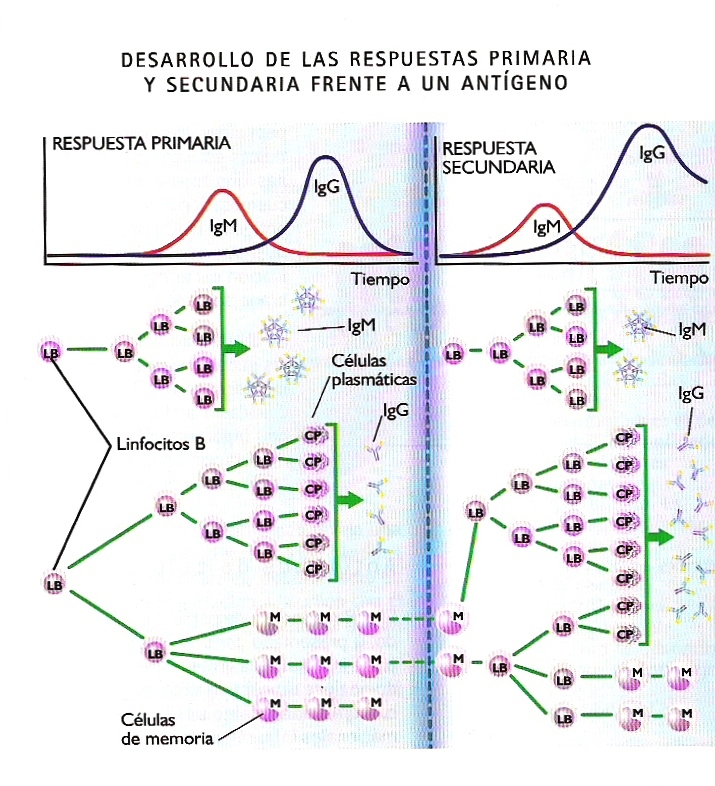
Los linfocitos son uno de los tipos celulares más abundantes en el organismo. Se calcula que en un ser humano adulto hay alrededor de un billón de linfocitos, lo que representa aproximadamente un 2% del peso corporal. En tal población de linfocitos, que se renueva a un ritmo de mil millones por día, hay células con receptores específicos para prácticamente cualquier antígeno posible. Cuando tiene lugar el primer contacto con el antígeno se desencadena una una respuesta primariahumoral y/o celular. Aquellos linfocitos B con receptores BCR capaces de reconocer el antígeno, con la ayuda de los linfocitos T auxiliares, se dividen intensamente y proliferan para dar lugar a un numeroso clon de células competentes para combatirlo. Lo mismo ocurre en su caso con los linfocitos T específicos del antígeno. En el clon seleccionado de linfocitos B pronto se diferencian dos líneas celulares: una de ellas, las denominadas células plasmáticas, se encarga de producir anticuerpos, inicialmente del isotipo IgM y a continuación del IgG. Tanto los anticuerpos circulantes como las células plasmáticas que los producen tienden a desaparecer gradualmente una vez neutralizado el antígeno, con lo que finaliza la respuesta primaria.
La otra línea celular la integran las denominadas células de memoria, que persisten durante largos períodos “acuarteladas” en los órganos linfáticos en prevención de un eventual nuevo contacto con el antígeno. Si ese contacto se produce, sobreviene la respuesta secundaria. Las células de memoria proliferan rápidamente dando lugar a un nuevo clon de células plasmáticas, mucho más numeroso que el generado en la respuesta primaria, que produce y libera una gran cantidad de anticuerpos del isotipo IgG. Todo ello ocurre en paralelo con un nuevo proceso de selección clonal similar al ocurrido durante la respuesta primaria.
En la respuesta secundaria se consigue una concentración de anticuerpos en los tejidos afectados mucho mayor que en la respuesta primaria, y además se consigue mucho más rápidamente, todo lo cual repercute positivamente en la eficacia de la respuesta (Figura 21.11).
{kind=link}
6.- INMUNIDAD NATURAL.
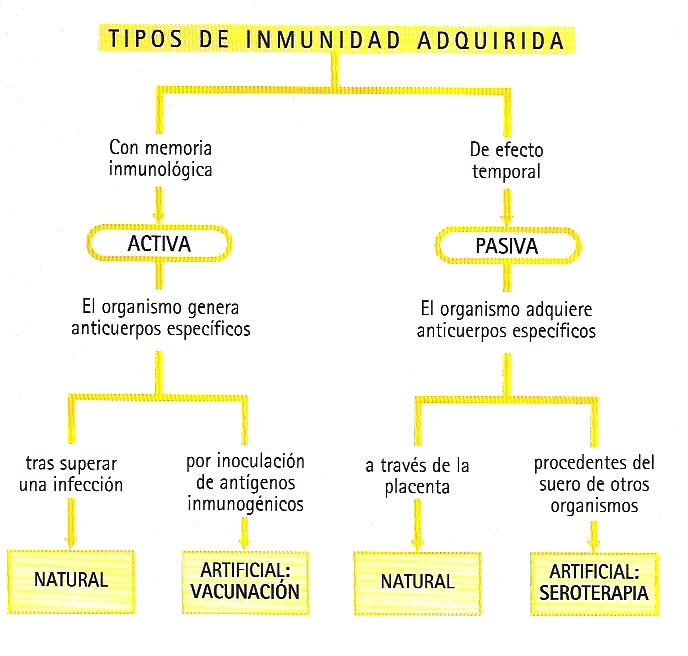
Además de utilizarse en sentido amplio para referirse al conjunto de mecanismos defensivos analizados en el presente capítulo, el término inmunidad se viene utilizando ya desde la antigüedad para aludir a un estado de resistencia que presenta un organismo frente a la infección. A la luz de los conocimientos actuales se dice que un organismo es inmune frente a un determinado germen cuando su sistema inmunitario es capaz de desactivarlo sin que llegue a aparecer una reacción patológica. La inmunidad frente a un determinado microorganismo patógeno puede ser congénita o adquirida.
{kind=link}
- Inmunidad congénita.- La confieren los mecanismos de defensa inespecíficos de las primeras líneas defensivas sin que medie contacto previo con el patógeno. Suele ser debida a características fisiológicas de la especie que la hacen incompatible con el microorganismo invasor. Así, por ejemplo, los pollos son inmunes frente al Bacillus antracis, agente causante del ántrax en distintas especies animales, debido a que su temperatura corporal es demasiado elevada para el crecimiento de este germen. También puede suceder que dentro de la misma especie determinados individuos presenten inmunidad congénita frente a una enfermedad debido a causas genéticas. Por ejemplo, se ha podido comprobar que aproximadamente un 1% de la humanidad presenta inmunidad congénita frente al VIH, el agente causante del SIDA, debido a causas genéticas todavía no aclaradas.
- Inmunidad adquirida.- La confieren los mecanismos de defensa específicos y está relacionada con el fenómeno de la memoria inmunitaria, anteriormente descrito. Es conocido ya desde la antigüedad que determinados gérmenes inducen en los individuos infectados resistencia frente a nuevas infecciones durante largos períodos. La explicación de este fenómeno estaría en las características de las respuestas inmunitarias primaria y secundaria y en la persistencia en los órganos linfáticos de células de memoria específicas del germen. El tipo de inmunidad descrito se adquiere de forma activa, pues es el propio organismo hospedador el que genera la respuesta inmunitaria frente al germen. La inmunidad natural también se puede adquirir de forma pasiva, mediante anticuerpos fabricados por otro organismo, como en el caso de los transmitidos de madre a hijo a través de la placenta o de la leche materna. La inmunidad natural pasiva, al depender de la persistencia de los anticuerpos suministrados y no de células de memoria, es siempre mucho menos duradera que la activa.
7.- INMUNIDAD ARTIFICIAL: SUEROS Y VACUNAS.
Desde los primeros tiempos de la ciencia inmunológica, los investigadores trataron de encontrar la manera de inducir artificialmente en seres humanos el estado de resistencia frente a la enfermedad que hemos descrito con el nombre de inmunidad. Se puede afirmar que los intentos de combatir la enfermedad por medio de la inmunidad artificial han constituido el auténtico hilo conductor de la inmunología durante los últimos 200 años. Los éxitos obtenidos en esta lucha representan una de las mayores contribuciones de la ciencia al bienestar de la humanidad.
Podemos distinguir entre dos formas de inmunidad artificial:
- INMUNIDAD ARTIFICIAL PASIVA.- Se induce mediante la inyección de sueros. Un suero es un preparado artificial que contiene anticuerpos contra el germen que se desea combatir. Estos anticuerpos han sido sintetizados por un organismo diferente del receptor previamente inoculado con dicho germen. Antiguamente se utilizaban animales para la obtención de sueros, pero en la actualidad se obtienen a partir de cultivos de células humanas, que producen inmunoglobulinas mucho más parecidas a las del organismo receptor.
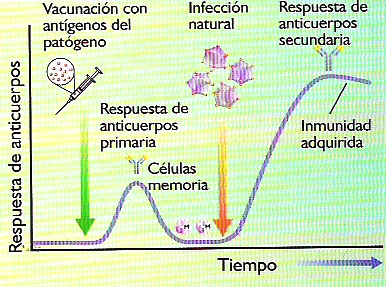
- INMUNIDAD ARTIFICIAL ACTIVA.- Se induce mediante la inyección de vacunas. Una vacuna es un preparado artificial que contiene antígenos característicos del germen que se desea combatir. La vacuna induce en el organismo receptor una respuesta inmunitaria primaria con producción de células de memoria. Si tras la vacunación se produce una infección natural por el mismo germen, se desencadena directamente una respuesta secundaria que la neutraliza con mucha más eficacia que en el caso de ausencia de vacuna (Figura 21.12). Así pues, la vacunación consigue emular el proceso natural que confiere inmunidad adquirida sin los inconvenientes que supone el padecer una primera infección y la consiguiente reacción patológica.
{kind=link}
Las vacunas pueden obtenerse a partir de microorganismos vivos atenuados, es decir, a los que se les ha reducido artificialmente su virulencia. Estas vacunas son muy eficaces pero conllevan cierto peligro de infección patológica, sobre todo en individuos con alguna inmunodeficiencia. Otra forma de obtener vacunas es a partir de microorganismos muertos o de fragmentos de microorganismos con capacidad antigénica. Estas vacunas tienen menor capacidad inmunogénica, por lo que son menos eficaces y requieren administrar dosis de recuerdo; sin embargo, resultan más seguras, ya que no pueden dar lugar a infección patológica.
8.- TRASTORNOS DEL SISTEMA INMUNITARIO.
Aunque el sistema inmunitario específico es, dada su extraordinaria capacidad para procesar y manejar información, uno de los productos más sofisticados de la evolución biológica, en ocasiones pueden producirse fallos en su funcionamiento, que repercuten en la capacidad del organismo afectado para hacer frente a los agentes externos. Analizaremos a continuación los distintos tipos de anomalías que pueden afectar al sistema inmunitario.
- INMUNODEFICIENCIAS.- Se trata de estados patológicos en los que el sistema inmunitario se encuentra total o parcialmente dañado de manera que no puede cumplir con sus funciones de defensa del organismo, con lo que éste resulta más vulnerable a las infecciones por agentes externos así como a procesos cancerosos. Las inmunodeficiencias pueden ser de dos tipos:
- Congénitas (o primarias).- Obedecen a defectos hereditarios, consecuencia de mutaciones que afectan a los genes responsables del funcionamiento del sistema inmunitario (genes que codifican las inmunoglobulinas o las proteínas implicadas en su síntesis, genes que codifican los receptores de los linfocitos B y T, y otros muchos). En general se manifiestan ya en las primeras etapas de la vida, dando lugar a síndromes como los de los llamados “niños burbuja”.
- Adquiridas (o secundarias).- Obedecen a causas externas al organismo que influyen en el desarrollo y funcionamiento de diversas estructuras relacionadas con el sistema inmunitario, como los distintos tipos de linfocitos o los órganos linfoides en los que aquéllos maduran. Una causa de este tipo de inmunodeficiencias puede ser una nutrición deficiente o escasa. Sin embargo, las inmunodeficiencias adquiridas más importantes se deben a la acción de agentes infecciosos que atacan selectivamente a las células del sistema inmunitario. Entre ellas destaca el síndrome de inmunodeficiencia adquirida (SIDA) producido por el VIH.
· HIPERSENSIBILIDADES.- Son estados patológicos que se producen como consecuencia de una respuesta inmunitaria de excesiva intensidad ante un antígeno de escasa peligrosidad para el organismo. Las reacciones de hipersensibilidad más comunes son conocidas como alergias. Se desencadenan ante una gran variedad de antígenos, que pueden ser de origen natural (granos de polen, heces de ácaros, diversas secreciones de los vegetales, etc.) o artificial (plásticos, metales, aditivos alimentarios, etc.). El trastorno generado por estas respuestas inmunitarias particularmente intensas es consecuencia de la reacción inflamatoria que las acompaña y sus efectos sobre los tejidos (edema, enrojecimiento, fiebre, picor, etc.).
Además de las alergias, existen reacciones de hipersensibilidad masivas que pueden acarrear la muerte del organismo afectado. Es el caso que se da a consecuencia de transfusiones de sangre entre un donante y un receptor con grupos sanguíneos incompatibles: el ataque de los anticuerpos del receptor contra los glóbulos rojos antigénicos del donante produce una coagulación masiva de la sangre (choque anafiláctico) de consecuencias fatales.
- AUTOINMUNIDAD.- Se dan cuando un organismo desencadena una respuesta inmunitaria contra moléculas propias denominadas autoantígenos. El origen de estos fenómenos está en un error del sistema inmunitario a la hora de identificar los antígenos. Las causas de estos errores pueden ser múltiples (mutaciones en los genes del MHC, similitudes entre la conformación tridimensional de algunas zonas del autoantígeno con los epitopos de un antígeno externo previamente identificado, y otras muchas). Por regla general, los fenómenos autoinmunes se manifiestan en forma de una respuesta inmunitaria celular, con ataque de los linfocitos T citotóxicos a una determinada estirpe de células del organismo afectado.
No hay comentarios:
Publicar un comentario