GENÉTICA MOLECULAR.
1.- INTRODUCCIÓN: EL CONCEPTO CLÁSICO DE GEN Y EL NACIMIENTO DE LA GENÉTICA MOLECULAR.
El redescubrimiento de los trabajos de Gregor Mendel en el año 1900 y la amplia generalización de sus conclusiones a la que condujeron los trabajos de genetistas como Morgan, Sturtevant y Muller en las primeras décadas del S. XX, trajeron como consecuencia la aceptación prácticamente universal de los principios mendelianos de la herencia biológica. Esta aceptación propició grandes avances en el conocimiento de los procesos genéticos que afectan tanto a las células individuales como a los organismos pluricelulares y a las poblaciones de seres vivos.
El gran bagaje de conocimientos acumulados, que globalmente configuran lo que se ha dado en llamar genética clásica, no sólo ha elevado nuestro nivel de comprensión de los sistemas vivos hasta cotas insospechadas pocos años antes, sino que su aplicación en campos como la agricultura y la medicina ha resultado enormemente beneficiosa para la humanidad. Sin embargo, durante todo este período, que abarca la primera mitad del S. XX, el concepto fundamental de la genética, el gen, permanecía desprovisto de todo contenido material. Aunque la teoría cromosómica de la herencia había establecido con claridad la localización de los genes en el núcleo celular y, más concretamente, en los cromosomas, los genetistas clásicos desconocían por completo la naturaleza físico-química del gen, así como los mecanismos por los que éste, desde su sede en el núcleo celular, era capaz de dirigir la maquinaria bioquímica de la célula y de replicarse con exactitud a lo largo de muchas generaciones celulares. El gen mendeliano era una entidad indivisible y abstracta cuya existencia era reconocida por sus efectos sobre células y organismos aunque su naturaleza material continuase siendo un misterio. A la pregunta "¿qué es un gen?" un genetista clásico probablemente respondiese que el gen es "algo" capaz de controlar un carácter hereditario, de replicarse a sí mismo fielmente en las sucesivas generaciones celulares, de recombinar con otros genes en el proceso de división celular meiótica y de cambiar globalmente su estructura para producir una alternativa diferente del carácter que controla en el proceso conocido como mutación. Conocer la naturaleza física del gen y los mecanismos moleculares mediante los cuales éste se replica y controla un carácter hereditario, aunque de indudable interés intelectual para el genetista clásico, no es el objetivo de su trabajo, puesto que las teorías y predicciones experimentales que formula acerca de los mecanismos de la herencia, y el éxito de las mismas, no dependen de estos conocimientos.
En los años 40, el panorama hasta aquí dibujado cambió radicalmente cuando un nutrido grupo de científicos, cuya formación y motivaciones eran muy diferentes de las de los genetistas clásicos, comenzó a interesarse por la naturaleza del gen. Se trataba de investigadores que se habían formado en el campo de las ciencias físicas y que estaban escasamente familiarizados con los conocimientos acumulados en las décadas precedentes por los genetistas clásicos e, incluso, con la Biología en general. Por razones que desarrollaremos a continuación, este grupo de científicos centró su interés en la resolución de un único problema: la base física de la información genética.
Los antecedentes de este movimiento intelectual hay que buscarlos en la exposición, por parte de Niels Bohr (Figura 19.1), uno de los más ilustres físicos del S. XX, de la idea de que algunos de los fenómenos biológicos podrían no ser completamente explicables en función de conceptos físicos y químicos convencionales. En opinión de Bohr, y de algunos de sus discípulos, la herencia biológica era claramente uno de estos fenómenos.
Las ideas de Bohr, que en algún momento fueron mal interpretadas y tergiversadas con la intención de resucitar la vieja doctrina filosófica del vitalismo, no llegaron a calar hondo entre la comunidad científica hasta que en 1945 (inmediatamente después del final de la segunda guerra mundial), Erwin Schrödinger (Figura 19.2), uno de los padres de la mecánica cuántica, publicó un pequeño ensayo titulado "¿Qué es la vida?", en el que dichas ideas eran recogidas y desarrolladas de manera mucho más rigurosa. Para Schrödinger, el único problema real, aquel en el que las explicaciones físicas convencionales podrían resultar insuficientes, era la naturaleza física del gen.
 Las estimaciones de los tamaños de los genes que se deducían de los análisis realizados por los genetistas clásicos en la mosca del vinagre (Drosophila melanogáster) indicaban que éstos eran similares a los de las mayores moléculas conocidas. Si, como apuntaban estas estimaciones, el gen no era más que un tipo particular de molécula, se trataba, en opinión de Schrödinger, de una molécula muy especial. En primer lugar, el gen demostraba ser una molécula altamente estable, capaz de conservar su estructura específica, y por lo tanto su contenido informativo, durante largos períodos de tiempo y en un ambiente químicamente heterogéneo como es el ambiente celular. En segundo lugar, lo que resultaba todavía más desconcertante, la "molécula génica" era capaz de dar lugar a copias fieles de sí misma y transmitirlas sin alteración a lo largo de innumerables generaciones celulares. No existía ninguna molécula conocida que reuniera estas características.
Las estimaciones de los tamaños de los genes que se deducían de los análisis realizados por los genetistas clásicos en la mosca del vinagre (Drosophila melanogáster) indicaban que éstos eran similares a los de las mayores moléculas conocidas. Si, como apuntaban estas estimaciones, el gen no era más que un tipo particular de molécula, se trataba, en opinión de Schrödinger, de una molécula muy especial. En primer lugar, el gen demostraba ser una molécula altamente estable, capaz de conservar su estructura específica, y por lo tanto su contenido informativo, durante largos períodos de tiempo y en un ambiente químicamente heterogéneo como es el ambiente celular. En segundo lugar, lo que resultaba todavía más desconcertante, la "molécula génica" era capaz de dar lugar a copias fieles de sí misma y transmitirlas sin alteración a lo largo de innumerables generaciones celulares. No existía ninguna molécula conocida que reuniera estas características.
Schrödinger sugería en su ensayo que la molécula génica podría ser un gran cristal aperiódico consistente en la sucesión de unos cuantos elementos isómeros y que la naturaleza exacta de esta sucesión constituiría el código genético. Apuntaba, además, que, por el hecho de que las propiedades exhibidas por la molécula génica no resultaran explicables desde el punto de vista de las leyes físicas conocidas hasta la fecha, no había que presuponer que dicha molécula eludiese dichas leyes. Por el contrario, estas propiedades podrían implicar la existencia de "otras leyes físicas", desconocidas por el momento, que, una vez descubiertas, formarían parte integral de esta ciencia junto con las ya conocidas.
La propuesta de Schrödinger tuvo efectos inmediatos. Los físicos de la época se encontraban sumidos en un gran malestar profesional, probablemente relacionado con el uso bélico que se había hecho de sus investigaciones al final de la segunda guerra mundial, y estaban deseosos de dirigir sus esfuerzos hacia nuevas fronteras del conocimiento. En este contexto, un físico del prestigio de Erwin Schrödinger expone la idea de que el estudio de la materia viva, y más concretamente del gen, podría revelar la existencia de "otras leyes físicas" que aguardaban ahí a que alguien las descubriese. Animados por esta posibilidad, un buen número de físicos decidió abandonar el campo de la investigación para el que habían sido formados y "desembarcaron" en las ciencias biológicas con el objetivo de esclarecer la base físico-química de la información genética.
2.- LA NATURALEZA DEL MATERIAL HEREDITARIO. 
En 1868, un médico alemán llamado Friedich Miescher (Figura 19.3) descubrió, cuando analizaba la composición de núcleos de células del pus procedente de vendajes quirúrgicos, un cuarto tipo de sustancia que se añadía a los ya por entonces conocidos glúcidos, lípidos y proteínas como componente esencial de la materia viva. Se trataba de una sustancia ácida, rica en fósforo, que Miescher denominó nucleína y que poco después empezó a conocerse con el nombre de ácido nucleico, denominación esta que hacía referencia tanto a su carácter ácido como a su localización en el núcleo celular.
Resulta paradójico que esta sustancia fuese descubierta sólo tres años después de que Mendel estableciese el concepto de gen y que hubiesen de transcurrir otros noventa hasta que dicha sustancia fuese reconocida universalmente como su soporte material.
El conocimiento preciso de la química elemental de los ácidos nucleicos se demoró bastante con respecto al de otras biomoléculas orgánicas, probablemente debido a su mayor complejidad estructural. En los primeros años del S. XX fueron identificados sus componentes moleculares (pentosas, bases nitrogenadas y ácido fosfórico). En la década de los años 20 se descubrió cómo se unen entre sí estos componentes para dar lugar a los nucleótidos y cómo se enlazan éstos para dar lugar a los ácidos nucleicos. Por la misma época se pudo reconocer la existencia de dos tipos principales de ácido nucleico (DNA y RNA) así como las diferencias estructurales entre ellos. Todavía a comienzos de los años 40 se desconocía el hecho de que los ácidos nucleicos eran en realidad macromoléculas que consistían en largas cadenas polinucleotídicas, formadas por centenares, miles e incluso millones de nucleótidos unidos por puentes fosfodiéster. Cuando en 1945 Schrödinger propone a sus colegas físicos investigar la naturaleza del material genético, era comúnmente aceptado que la molécula de DNA consistía en un tetranucleótido cíclico, formado por los cuatro desoxirribonucleótidos conocidos. La idea del tetranucleótido provenía de los análisis cuantitativos realizados sobre DNAs de unas pocas especies de seres vivos, que indicaban que en todos ellos las 4 bases nitrogenadas (A, T, G y C) se encontraban en cantidades iguales.
Por otra parte, en la teoría cromosómica de la herencia se establecía ya la localización de los genes a lo largo de los cromosomas, dentro del núcleo celular. El análisis químico de los cromosomas indicaba que éstos estaban compuestos por DNA y proteínas a partes aproximadamente iguales. Todo indicaba pues que alguna de estas dos sustancias debía ser el material hereditario.
La química elemental de las proteínas era bien conocida ya cuando empezó a plantearse el problema de la naturaleza del gen. Aunque sólo se habían dado los primeros pasos en la determinación de su estructura tridimensional, se había establecido ya con claridad que las moléculas proteicas consistían en largas cadenas de aminoácidos unidos mediante enlaces peptídicos. También se había puesto de manifiesto que la identidad química de cada molécula proteica vendría dada por la naturaleza y posición de los diferentes restos de aminoácidos a lo largo de la cadena polipeptídica, es decir, por su estructura primaria.
En este contexto histórico, con un conocimiento bastante avanzado de la estructura química de las proteínas y relativamente pobre de la de los ácido nucleicos, un buen número de investigadores se decantó inicialmente por las proteínas como principales candidatas a constituir la base química de la herencia. Resultaba claro para ellos que las largas cadenas de aminoácidos de las proteínas respondían mejor a la idea del gran cristal aperiódico formulada por Schrödinger que el pequeño tetranucleótido cíclico en que parecía consistir la molécula de DNA. Según su punto de vista, la información genética podría estar cifrada en forma de diferentes secuencias de aminoácidos, siendo éstos los "elementos isómeros" a los que Schrödinger hacía referencia. En los próximos apartados de este capítulo comprobaremos que el tiempo y los hechos vinieron a quitarles la razón.
2.1.- EL EXPERIMENTO DE AVERY.
 La primera prueba de que el DNA era después de todo el material genético, fue obtenida en 1944 por Oswald T. Avery, y sus colaboradores C.M. McLeod y M.J. McCarty en el transcurso de un trabajo experimental en el que trataban de encontrar explicación a un fenómeno observado algunos años antes por el médico británico F. Griffith (Figura 19.4).
La primera prueba de que el DNA era después de todo el material genético, fue obtenida en 1944 por Oswald T. Avery, y sus colaboradores C.M. McLeod y M.J. McCarty en el transcurso de un trabajo experimental en el que trataban de encontrar explicación a un fenómeno observado algunos años antes por el médico británico F. Griffith (Figura 19.4).
En 1928 F. Griffith estudiaba el proceso de infección en ratones porStreptococcus pneumoniae, más conocido como "neumococo", una bacteria que se encuentra entre los agentes causantes de la neumonía humana y que resulta especialmente patógena para el ratón: la inyección en un ratón de esputos procedentes de un paciente afectado de neumonía neumocócica le ocasiona a aquél la muerte en menos de 24 horas. El neumococo debe su carácter patógeno a una cápsula de polisacáridos que lo protege de los mecanismos de defensa del animal infectado. Griffith había aislado una cepa mutante de esta especie, que había perdido su capacidad para sintetizar esta cápsula y que resultaban por lo tanto vulnerables a dichos mecanismos de defensa: los ratones inoculados con bacterias de esta cepa no contraían la neumonía y por consiguiente sobrevivían. Ambas variantes podían distinguirse una de la otra con facilidad debido al aspecto de las colonias que formaban en las placas de cultivo, que tenían aspecto brillante (S) en la variante patógena común y aspecto rugoso (R) en la variante mutante no patógena. El aspecto brillante o rugoso de las colonias era también una consecuencia de la presencia o ausencia respectivamente de la mencionada cápsula de polisacáridos.
En el curso de sus investigaciones Griffith descubrió con sorpresa que los ratones inoculados con mutantes R no patógenos mezclados con una muestra de bacterias S patógenas previamente muertas por efecto del calor, contraían la neumonía y morían a las pocas horas. Las bacterias recuperadas de la sangre de los ratones muertos habían recuperado su capacidad para sintetizar la cápsula de polisacáridos y con ello su carácter patógeno y el aspecto brillante de las colonias a las que daban lugar. El contacto con las bacterias S había producido en las bacterias R una transformación RàS que se transmitía a las sucesivas generaciones celulares. Años más tarde Griffith comprobó que no era imprescindible que el contacto entre las dos cepas bacterianas se produjese en el interior del ratón: la transformación también se producía en cultivos de bacterias R que crecían en contacto con bacterias S muertas. Más significativo aún resultó el hecho de que la transformación se produjese como consecuencia del contacto de cultivos de bacterias R creciendo en contacto con un"extracto libre de células" de bacterias S, es decir, no era imprescindible la estructura celular intacta de las bacterias S muertas sino que una disolución de sus componentes moleculares solubles era suficiente.

Los experimentos de Griffith fueron el punto de partida del trabajo de Avery, McLeod y McCarthy, que se plantearon identificar, en el extracto libre de células que se ha mencionado, la naturaleza química del "principio transformante"responsable del fenómeno observado. Para ello llevaron a cabo un fraccionamiento sistemático del extracto libre de células y ensayaron la capacidad transformante de las distintas fracciones sobre cultivos de bacterias R. Tras ensayar con distintas fracciones del extracto (lípidos, glúcidos, proteínas, etc.) con resultados negativos, comprobaron que la fracción que contenía los ácidos nucleicos inducía eficazmente la transformación. Un fraccionamiento ulterior llevó a la conclusión de que el principio transformante buscado no era otro que el DNA bacteriano: pequeñísimas cantidades de este DNA purificado eran suficientes para transformar las bacterias R en bacterias S. Avery y sus colaboradores demostraron también que el DNA de las bacterias transformadas y de sus descendientes podía a su vez inducir la transformación en otras bacterias R y que en sucesivos ciclos de transformación como los descritos se mantenía esta capacidad.
Tras estas experiencias las conclusiones de Avery (Figura 19.6) estaban cada vez más claras: el DNA de las bacterias S muertas era la sustancia que contenía la información necesaria para hacer que las bacterias R y su descendencia recuperasen su capacidad para sintetizar su cápsula de polisacáridos y con ella su carácter patógeno, es decir, el DNA era el material genético de Streptococcus pneumoniae.
La publicación de los resultados de Avery, McLeod y McCarthy en 1944 provocó una oleada de escepticismo crítico entre la comunidad científica de la época. Como ya se ha dicho, la mayoría de los investigadores apuntaba a las proteínas como principales candidatas a constituir la base química del gen. Las principales objeciones incidían en el hecho de que las técnicas al uso de fraccionamiento y purificación de macromoléculas no eran eficaces al 100%, de manera que una pequeña cantidad de proteínas contaminando el extracto de DNA purificado pudiera ser la responsable de la actividad transformante de éste. Analizada desde la perspectiva actual esta objeción parece claramente formulada ad hoc en defensa de una idea previa demasiado firmemente asentada: “tenían que ser las proteínas”. En efecto, si el material genético consiste en proteínas, no parece lógico afirmar que la actividad transformante reside en una mínima cantidad de proteína contaminante en la fracción de DNA purificado y no en la que contiene la mayor parte de las proteínas de la célula. Sin embargo, tales críticas propiciaron la realización de nuevos controles experimentales que siempre confirmaron las conclusiones precedentes. El equipo de Avery trató el extracto el DNA purificado de las bacterias S con proteasas (enzimas que degradan las proteínas) sin que esto afectara a su actividad transformante. Por otra parte, el tratamiento con desoxirribonucleasas (enzimas que degradan el DNA) destruía en pocos minutos cualquier rastro de dicha actividad. Otros experimentos, realizados por R.D. Hotchkiss, confirmaron que la actividad transformante del DNA no se restringía al carácter virulento o no de las cepas bacterianas, ni al aspecto liso o rugoso de sus colonias, sino que también operaba de manera análoga para otros caracteres hereditarios, como la resistencia a distintos tipos de antibióticos.
Los nuevos resultados experimentales no consiguieron diluir el escepticismo reinante. Si nueve años después de la publicación de los trabajos de Avery la comunidad científica aceptó por fin el papel del DNA como molécula portadora de la información genética, no fue por que se hubiesen realizado nuevos controles con resultados más convincentes, sino porque los grandes avances que se produjeron en este período acerca de la química de los ácidos nucleicos disiparon cualquier duda al respecto.
2.2.- LA REGLA DE EQUIVALENCIA DE CHARGAFF.
La principal dificultad a la hora de reconocer al DNA un papel relevante en la herencia biológica residía en que nadie comprendía, a la luz de los conocimientos existentes acerca de la química de los ácidos nucleicos, cómo podía desempeñarlo. Ya se ha comentado que a comienzos de los años cuarenta se consideraba que la molécula de DNA era un tetranucleótido cíclico formado por los cuatro desoxirribonucleótidos posibles (dAMP, dGMP, dTMP y dCMP). Incluso cuando en el curso de esta década se estableció que el DNA era en realidad una macromolécula con peso molecular muy superior al que cabría esperar de un simple tetranucleótido, se interpretó que éste vendría a ser la unidad monomérica repetitiva que formaba los grandes polímeros de DNA, de manera similar a lo que representaba la glucosa en los polímeros monótonos de almidón y glucógeno. Era evidente que una macromolécula con estas características no podía ser el cristal aperiódico de Schrödinger.
 La persistencia de la idea del tetranucleótido tenía su razón de ser en que los análisis de la composición química de DNA procedente de diferentes fuentes biológicas parecían arrojar siempre proporciones equimolares de las cuatro bases nitrogenadas (A, G, T y C). A finales de los años 40 Erwin Chargaff (Figura 19.7) adaptó las recién descubiertas técnicas de cromatografía sobre papel a la separación y cuantificación de los componentes de los ácidos nucleicos y las empleó en el análisis de diferentes muestras de DNA. La mayor precisión de estas técnicas le permitió comprobar que las cuatro bases nitrogenadas no se encontraban necesariamente en proporciones exactamente iguales. Además, Chargaff encontró que la composición en bases nitrogenadas difiere ampliamente según la procedencia biológica de la muestra de DNA.
La persistencia de la idea del tetranucleótido tenía su razón de ser en que los análisis de la composición química de DNA procedente de diferentes fuentes biológicas parecían arrojar siempre proporciones equimolares de las cuatro bases nitrogenadas (A, G, T y C). A finales de los años 40 Erwin Chargaff (Figura 19.7) adaptó las recién descubiertas técnicas de cromatografía sobre papel a la separación y cuantificación de los componentes de los ácidos nucleicos y las empleó en el análisis de diferentes muestras de DNA. La mayor precisión de estas técnicas le permitió comprobar que las cuatro bases nitrogenadas no se encontraban necesariamente en proporciones exactamente iguales. Además, Chargaff encontró que la composición en bases nitrogenadas difiere ampliamente según la procedencia biológica de la muestra de DNA.
Puede resultar hoy extraño el que no se detectase antes esta amplia variación. Lo cierto es que en las muestras de DNA analizadas hasta entonces, la mayoría de ellas procedentes de organismos eucariontes, las proporciones de las distintas bases nitrogenadas oscilaban unos pocos puntos porcentuales con respecto al 25% que exigía la hipótesis del tetranucleótido, de manera que los primitivos métodos de análisis no permitían distinguir los resultados de los que se obtendrían si efectivamente las cuatro bases se encontrasen en proporciones equimolares. En los años subsiguientes se analizaron muestras de DNA procedentes de diferentes especies bacterianas, pudiendo comprobarse que en las células procariotas el espectro de variación de la composición en bases nitrogenadas es todavía mucho más amplio que en las células eucariotas.
La publicación de los resultados de Chargaff no sólo trajo consigo el rechazo definitivo de la hipótesis del tetranucleótido, sino que provocó un vuelco en la opinión de los científicos acerca del papel de los ácidos nucleicos. En efecto, si el DNA no era después de todo un polímero monótono, se había esfumado el principal inconveniente de la candidatura de esta macromolécula a constituir la base físico-química de la herencia biológica. Un largo polímero formado por cuatro tipos de nucleótidos en diferentes ordenaciones sí podía ser el cristal aperiódico formado por unos pocoselementos isómeros que Schrödinger había sugerido. Además, el hecho de que la composición en bases nitrogenadas del DNA variase ampliamente de unas especies a otras podía ser un reflejo de su especificidad biológica. La información genética podría muy bien estar cifrada en forma de la secuencia específica de bases nitrogenadas de la cadena polinucleotídica y el fenómeno de la mutación podría explicarse como un cambio fortuito en dicha secuencia.
Los analistas de la historia de la Biología molecular dudan a la hora de atribuir a un científico en particular la formulación de las ideas precedentes. Más bien se inclinan por afirmar que, a partir de 1950, la teoría parecía “flotar en el ambiente”, provocando una gran efervescencia investigadora alrededor de la molécula de DNA en los años sucesivos.
Además de todo lo dicho hasta ahora, los resultados publicados por Chargaff contenían una información adicional acerca de la molécula de DNA, que andando el tiempo resultó de importancia capital: a pesar de la amplia variación encontrada entre las muestras de DNA de diferentes especies, en todas ellas la proporción molar entre el total de bases púricas y el de bases pirimídicas era próxima a 1. Lo mismo parecía ocurrir con la adenina y la timina por una parte y la guanina y la citosina por otra. Esta afirmación, conocida como la “regla de equivalencia de Chargaff”, revela un rasgo esencial de la molécula de DNA: aunque la composición en bases nitrogenadas de este polímero puede variar sin ninguna restricción, se cumple siempre que el número de adeninas es igual al de timinas y el de guaninas igual al de citosinas; y, como corolario, que el número de bases púricas es igual al de bases pirimídicas.
[A] = [T]
[G] = [C]
-----------------------------------
[A+G] = [T+C]
Aunque cuando publicó sus resultados Chargaff manifestó que la constancia de estas proporciones era algo “digno de resaltar” (“noticeable”), se mostró extraordinariamente prudente a la hora de considerar si se trataba de una simple coincidencia o si realmente revelaba algún rasgo importante de la molécula de DNA. Realmente, hubiera resultado asombroso que Chargaff pudiera intuir el significado de su regla de equivalencia con la información de que disponía en 1950.
2.3.- EL EXPERIMENTO DE HERSHEY Y CHASE.
Entre los microorganismos cuyo estudio contribuyó en mayor medida a la comprensión de la estructura y función del gen hay que destacar a los virus, que serán analizados en detalle en un capítulo posterior. A comienzos de los años 50 del S. XX los virus eran ya reconocidos como parásitos intracelulares obligados, con un grado de organización inferior al celular, que se reproducían dentro de las células a las que parasitaban. Se les relacionaba acertadamente con un buen número de enfermedades humanas de las que eran responsables y se había detectado su presencia en distintas especies animales y vegetales. También se habían descubierto virus que parasitaban y destruían células bacterianas, los llamados bacteriófagos.
La estructura de los virus es extremadamente simple en comparación con la de los organismos con organización celular: los más simples constan de una cápside de naturaleza proteica en la que se encierra un ácido nucleico que puede ser DNA o RNA.
 En 1952 Alfred D. Hershey y Marta Chase (Figura 19.8) diseñaron un experimento con el objeto de elucidar los detalles del proceso de infección de células bacterianas de la especieEscherichia Coli por bacteriófagos T4 (Figura 19.8b). Las partículas infecciosas de este fago están compuestas exclusivamente por DNA y proteínas. Hershey y Chase querían saber como se comportaba uno y otro tipo de macromoléculas durante el proceso de infección. Para ello, idearon una ingeniosa técnica de marcaje mediante isótopos radiactivos. Se percataron de que en las partículas virales la práctica totalidad de los átomos de fósforo se encontraban en el DNA (en los grupos fosfato de la cadena polinucleotídica) mientras que la práctica totalidad de los átomos de azufre se encontraban en las proteínas (en los aminoácidos metionina y cisteína). Así, decidieron utilizar los isótopos radiactivos 32P y 35S para delatar respectivamente la presencia de DNA y de proteínas. Para obtener partículas víricas marcadas permitieron el crecimiento de un cultivo de fagos T4 sobre células de E. coli en un medio en el que la única fuente de fósforo eran iones fosfato (PO43-) marcado con 32P, de manera que este isótopo se incorporaba a todas las biomoléculas de las células bacterianas y de los fagos que se reproducían en su interior. Por otra parte, hicieron lo propio con fagos obtenidos en un cultivo con iones sulfato (SO42-) marcado con 35S, que se incorporaría igualmente a las biomoléculas de bacterias y fagos. De este modo, una vez aislados y purificados los fagos obtenidos en uno y otro cultivo, dispusieron de dos cepas virales, una de ellas con el DNA marcado con 32P y otra con las proteínas marcadas con 35S.
En 1952 Alfred D. Hershey y Marta Chase (Figura 19.8) diseñaron un experimento con el objeto de elucidar los detalles del proceso de infección de células bacterianas de la especieEscherichia Coli por bacteriófagos T4 (Figura 19.8b). Las partículas infecciosas de este fago están compuestas exclusivamente por DNA y proteínas. Hershey y Chase querían saber como se comportaba uno y otro tipo de macromoléculas durante el proceso de infección. Para ello, idearon una ingeniosa técnica de marcaje mediante isótopos radiactivos. Se percataron de que en las partículas virales la práctica totalidad de los átomos de fósforo se encontraban en el DNA (en los grupos fosfato de la cadena polinucleotídica) mientras que la práctica totalidad de los átomos de azufre se encontraban en las proteínas (en los aminoácidos metionina y cisteína). Así, decidieron utilizar los isótopos radiactivos 32P y 35S para delatar respectivamente la presencia de DNA y de proteínas. Para obtener partículas víricas marcadas permitieron el crecimiento de un cultivo de fagos T4 sobre células de E. coli en un medio en el que la única fuente de fósforo eran iones fosfato (PO43-) marcado con 32P, de manera que este isótopo se incorporaba a todas las biomoléculas de las células bacterianas y de los fagos que se reproducían en su interior. Por otra parte, hicieron lo propio con fagos obtenidos en un cultivo con iones sulfato (SO42-) marcado con 35S, que se incorporaría igualmente a las biomoléculas de bacterias y fagos. De este modo, una vez aislados y purificados los fagos obtenidos en uno y otro cultivo, dispusieron de dos cepas virales, una de ellas con el DNA marcado con 32P y otra con las proteínas marcadas con 35S.
Con las cepas virales obtenidas Hershey y Chase procedieron a infectar dos cultivos de E. coli no marcados radiactivamente (cada uno de ellos con una cepa diferente). Tras permitir la infección por un corto período de tiempo separaron por centrifugación las partículas víricas que no habían conseguido adherirse a ninguna célula y volvieron a suspender las bacterias infectadas en un medio de cultivo nuevo. A continuación sometieron esta suspensión de bacterias infectadas a una violenta agitación por medio de un agitador Waring (un dispositivo que genera fuertes turbulencias en el líquido sobre el que actúa), de manera que las partículas víricas se desprendían (eran literalmente arrancadas) de la superficie celular sobre la que se habían fijado. Se procedió entonces a centrifugar la suspensión con el objeto de separar las bacterias, que se depositaban en el fondo del tubo de la centrífuga, de las partículas víricas sueltas, que permanecían en el líquido sobrenadante. Seguidamente, se midió en un contador Geiger la fracción total de radiactividad que se había depositado en el fondo del tubo y la que permanecía en el líquido sobrenadante. Por último, se ensayó la capacidad de las bacterias infectadas para producir nuevos fagos descendientes en su interior. Los resultados obtenidos fueron los siguientes:
- En el cultivo bacteriano infectado con fagos marcados con 32P, es decir, con DNA marcado radiactivamente, la mayor parte de la radiactividad se había depositado en el fondo del tubo de la centrífuga. La fracción que no lo había hecho así se encontraba en el líquido sobrenadante de la primera centrifugación, es decir, que se encontraba en las partículas víricas que no habían conseguido infectar a ninguna célula.
- En el cultivo bacteriano infectado con fagos marcados con 35S, es decir, con proteínas marcadas radiactivamente, la mayor parte de la radiactividad permanecía en el líquido sobrenadante obtenido tras la agitación violenta. La fracción restante correspondía a estructuras de la cápside viral que no se habían desprendido de la superficie celular por estar demasiado intensamente ligadas a ella.
- En ambos cultivos las bacterias infectadas recuperadas tras la agitación violenta conservaban prácticamente intacta su capacidad para dar lugar a nuevas progenies virales a su vez con capacidad infectiva.
Hershey y Chase extrajeron rápidamente conclusiones de estos resultados (Figura 19.8b. Hacer click para ver aumentado). De los dos componentes de la partícula vírica sólo el DNA penetraba en el interior de la célula durante el proceso de infección (por eso el 32P aparecía asociado a la fracción celular tras la centrifugación). Las proteínas del fago permanecían en el exterior de la célula durante todo el proceso de infección y se desprendían de la superficie celular por agitación (por eso el 35S aparecía en el líquido sobrenadante). Es decir, la partícula vírica infecciosa se fija a la superficie celular y de alguna manera “inyecta” su DNA en el interior de la célula. La cápside proteica, una vez inyectado en la bacteria el DNA que albergaba en su interior, ya no resulta más necesaria en el proceso de infección, como prueba el hecho de que la eliminación de estos “fagos fantasma” por agitación no altere la capacidad de las células infectadas para dar lugar a nuevas progenies virales. Es la molécula de DNA vírico la que, una vez dentro de la célula, parece tomar el control de su metabolismo para que éste se ponga al servicio del virus y comience a fabricar nuevas partículas infecciosas atendiendo a las instrucciones cifradas en esa misma molécula. En otras palabras: el DNA es el material genético del bacteriófago T4.
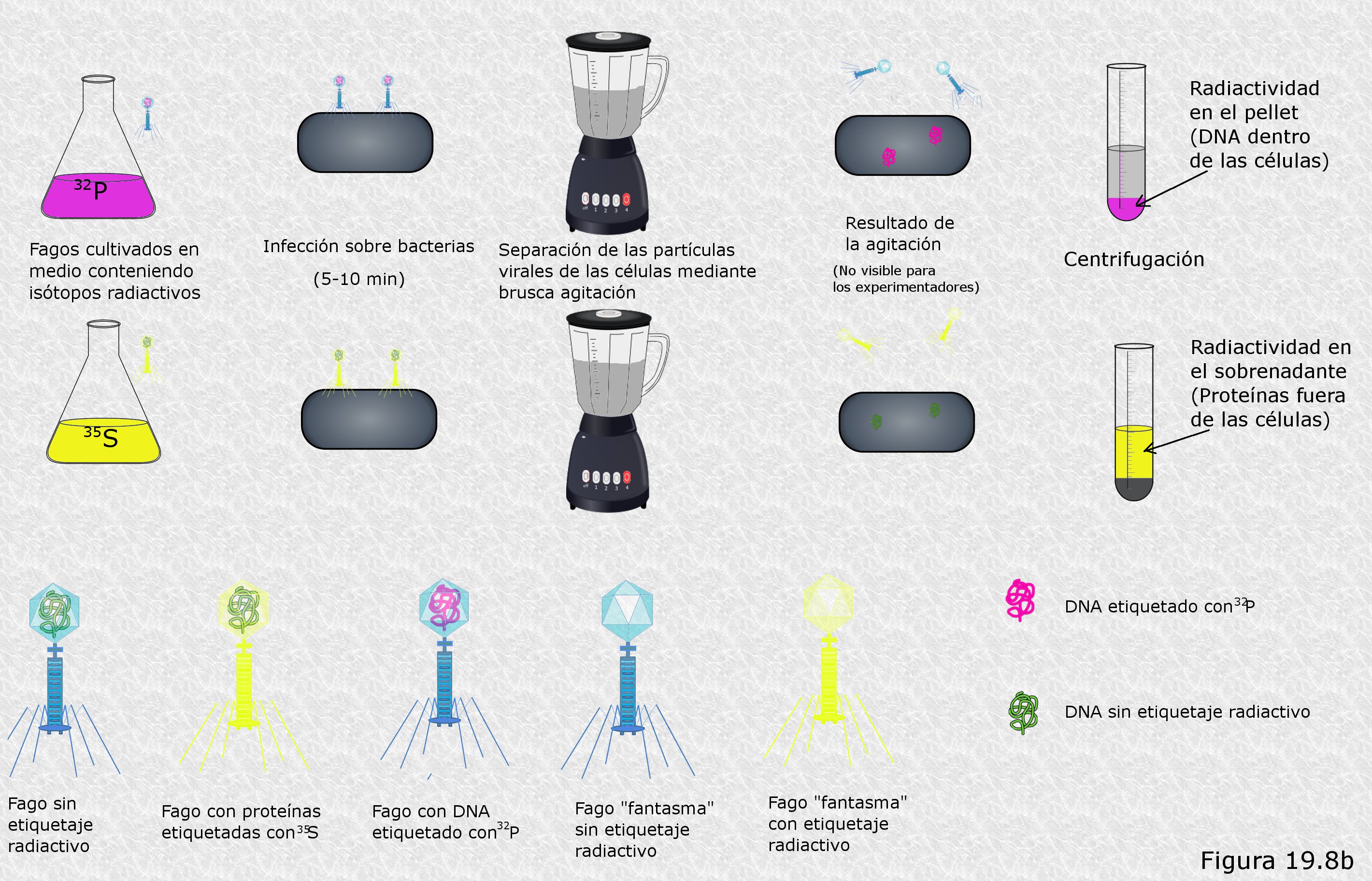
Los resultados obtenidos por Hershey y Chase eran especialmente concluyentes en lo que se refiere al papel del DNA como molécula de la herencia, pues en su experimento queda claro que el único nexo material entre dos generaciones sucesivas de bacteriófagos es una simple molécula de DNA.
Por otra parte, las conclusiones de este experimento concordaban y reforzaban las obtenidas ocho años antes por el equipo de Avery. Pero ahora el escenario había cambiado: los recientes descubrimientos acerca de la química de los ácidos nucleicos junto con los resultados obtenidos por Chargaff sobre la composición en bases nitrogenadas de diferentes muestras de DNA habían preparado el terreno para que estas conclusiones gozaran de una aceptación mucho mayor que la que se deparó a las publicadas por Avery. Probablemente, la publicación del trabajo de Hershey y Chase en el otoño de 1952 sirvió de estímulo para que un buen número de investigadores se concentraran durante los meses siguientes en elucidar la estructura tridimensional de la molécula de DNA.
No hay comentarios:
Publicar un comentario