Citogenética
cromómero (del griego khróma, color y meros, parte) es un disco coloreable, que se presenta en varias zonas a lo largo de un cromosoma.1 Cada cromómero contiene un número más o menos elevado de genes. Las bandas oscuras corresponden frecuentemente a cromatina inactiva.

{kind=link}
Los cromómeros alternan con zonas que no fijan los colorantes. Los cromómeros, son pequeños engrosamientos o nudosidades a manera de cuentas de collar, gracias a esto, los cromosomas se hacen visibles.
Éste bandeo es reproducible de núcleo a núcleo, formando un patrón constante de tal manera que los cromosomas pueden ser identificados y mapeados en toda su longitud. El patrón de bandeo que presentan los cromosomas politénicos es un reflejo constante de las secuencias de ADN, y por tanto las bandas sirven como marcadores para localizar varias características genéticas.
Se pueden visualizar durante las fases de la mitosis o de la meiosis de menor condensación de la cromatina (profase). Se hacen visibles en un cromosoma durante la profase de la meiosis en un subperiodo llamado Leptonema.
En el maíz hay más de 2.000 cromómeros en sus 20 cromosomas.Hay aproximadamente 5000 bandas y 5000 interbandas en total en el genoma de Drosophila melanogaster.
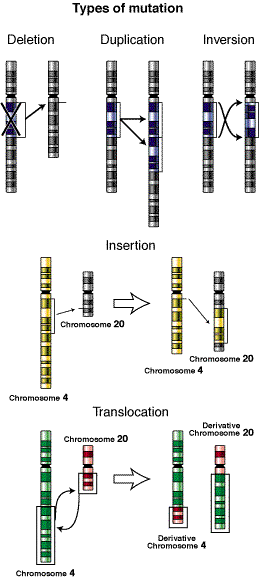
deleción, en genética, es un tipo especial de anomalía estructural cromosómica que consiste en la pérdida de un fragmento de ADN de un cromosoma. Esta pérdida origina un desequilibrio, por lo que las deleciones están incluidas dentro de las reordenaciones estructurales desequilibradas. El portador de una deleción es monosómico respecto a la información génica del segmento correspondiente del homólogo normal, por eso en ocasiones las deleciones son denominadas monosomías parciales.1
El origen de las deleciones puede ser una sencilla rotura cromosómica y pérdida del segmento acéntrico. En ciertos casos, las deleciones son el resultado de un entrecruzamiento desigual entre cromosomas homólogos o cromátidas hermanas mal alineados. También se pueden producir en la descendencia por segregación anormal de una translocación o una inversión equilibradas de los progenitores.
Tipos de deleciones
Las deleciones pueden ocurrir en cualquier parte del cromosoma. Según donde tenga lugar la pérdida del material genético se pueden clasificar en:
- Deleción proximal: El segmento de ADN ausente se encuentra cerca del centrómero.
- Deleción distal: La pérdida del material genético tiene lugar cerca de los telómeros.
Según el número de puntos de rotura en el cromosoma:
- Deleción terminal: Se produce solo una rotura en el cromosoma y todo el material genético se pierde desde la misma.
- Deleción intersticial: Hay dos puntos de rotura en el brazo del cromosoma. El material genético entre los mismo está ausente y las partes restante del cromosoma se fusionan provocando un acortamiento del brazo pero con el mismo principio y fin.
Algunas deleciones son tan pequeñas que no pueden ser apreciadas al microscopio óptico y son denominadas microdeleciones.
Incidencia
Las deleciones autosómicas citogenéticamente visibles tienen una incidencia de aproximadamente 1/7.000 nacidos vivos. Las deleciones submicroscópicas más pequeñas detectadas mediante análisis con micromatrices son mucho más frecuentes, si bien aún no se ha determinado el significado clínico de estas microdeleciones.1
Detección
Para detectar una deleción demasiado pequeña como para ser detectada en extensiones metafásicas se suelen utilizar las técnicas de bandeo de alta resolución o la técnica FISH. Para ser detectada por bandeo de alta resolución la deleción ha de ser de al menos varios millones de pares de bases. En cambio, la técnica FISH y las técnicas de micromatrices pueden detectar deleciones cariotípicamente indetectables o deleciones dudosas con consecuencias fenotípicas utilizando para ello sondas específicas de la región de interés.1
Consecuencias
Como consecuencia clínica se refleja la haploinsuficiencia, es decir, la incapacidad de la copia única del material genético para llevar a cabo las funciones que normalmente efectúan las dos copias. Las consecuencias parecen depender del tamaño del segmento delecionado y del número y funciones de los genes que contiene la deleción de material genético puede afectar desde un solo nucleótido (deleción puntual) a grandes regiones visibles citogenéticamente.1
La deleción de un gen o de parte de un gen puede ocasionar una enfermedad o una anomalía. Entre éstas destacan las siguientes:
- Síndrome del maullido del gato (deleción del brazo corto del cromosoma 5)
- Síndrome de Prader-Willi (deleción del brazo largo del cromosoma 15)
- Síndrome de Angelman (deleción de una región del cromosoma 15)
- Síndrome deleción 22q13 (deleción del extremo distal del brazo largo del cromosoma 22)
Deleciones
Un individuo es portador de una deleción cuando le falta un segmento cromosómico, si este segmento es un extremo del cromosoma, la alteración se denomina deficiencia. Si la deleción es muy grande es visible al microscopio óptico ya que el cromosoma presenta menor tamaño del normal.
La deleción en homocigosis suele ser letal para el individuo portador, si se presenta en heterocigosis, el efecto será más o menos deletéreo dependiendo de la importancia de los genes presentes en el segmento perdido. En individuos con determinación sexual XX-XY o XX-X0, las deleciones del cromosoma X son letales en los machos; En las hembras dependiendo del sistema de compensación de dosis génica, puede producir algunos efectos fenotípicos en el individuo heterocigótico. En la especie humana, en nacidos vivos, la deleción más frecuente y estudiada, es la conocida como síndrome de "Grito de gato", consiste en una deficiencia del brazo corto del cromosoma 5, que produce un retraso mental y finalmente la muerte del individuo.
En meiosis la configuración crítica para detectar una deleción es ver un bivalente heteromorfo, o bien observar una falta de apareamiento (bucle o lazo en el cromosoma no delecionado) en un segmento intersticial.
Dada la letalidad y el desequilibrio orgánico y cromosómico que producen las deleciones, la selección natural tiende a eliminarlas y por ello la importancia evolutiva de las deleciones es prácticamente nula.
 |  | 
duplicación cromosómica es la repetición de un fragmento de cromosoma a continuación del fragmento original. Las duplicaciones surgen por error en la duplicación del ADN, como producto de una reorganización cromosómica de tipo estructural o relacionado con un proceso de sobrecruzamiento defectuoso. Las duplicaciones no suelen serdeletéreas, son una fuente de nuevo material genético y base para nuevos cambios evolutivos. Muchas de las familias génicas con un origen evolutivo común, o las familias multigénicas pueden tener su origen en las duplicaciones. Si el segmento afectado es de gran tamaño, se puede detectar en meiosis con los mismos criterios que en las deleciones (bivalente heteromorfo o zona intersticial desapareada en el cromosoma con la duplicación).
Las duplicaciones no suelen tener una manifestación fenotípica observable a simple vista, sino mediante análisis citogenéticos y moleculares.
Papel de las duplicaciones en la evolución
Uno de los aspectos integrantes del estudio de la evolución es especular sobre los mecanismos posibles de la variación genética. En 1970, Susumo Ohno publico el polémico libro Evolution by Gene Duplication. La tesis de Ohno se basaba en la suposición de que los productos de genes esenciales son indispensables para la supervivencia de los miembros de cualquier especie a lo largo de la evolución. Estos genes no pueden acumular mutaciones que alteren su función primaria y dar lugar potencialmente a nuevos genes.1 Sin embargo, si se duplicara un gen esencial en una línea germina, en la copia extra se tolerarían cambios mutacionales proporciona la información genética para su función esencial. La copia duplicada quedaría libre para adquirir muchos cambios mutacionales durante largos periodos de tiempo. En periodos cortos, la nueva información genética podría no tener ventajas prácticas. Sin embargo, en periodos evolutivos largos, el gen duplicado podría cambiar lo suficiente como para que su producto asumiera un papel divergente en la célula. La nueva función podría dar una ventaja “adaptativa” al organismo, incrementando su eficacia biología. Ohno ha imaginado un mecanismo mediante el cual pudo haberse originado sustancial variabilidad genética. La tesis de Ohno está apoyada por el descubrimiento de genes que tiene una parte importante de sus secuencias de ADN en común, pero cuyos productos génicos son distintos.
La importancia evolutiva de las duplicaciones radica en el hecho de que los individuos portadores tienen dos copias de un mismo gen. En un individuo normal una mutación de ese gen puede tener efectos deletéreos, pero si hay dos copias y se produce una mutación en una de ellas, el individuos podrá seguir manifestando un fenotipo "aparentemente normal" y la selección natural no actuaría en su contra. Mediante este proceso se pueden ir originando nuevas copias de un mismo gen y producirse variantes y alternativas no alélicas a una secuencia de ADN. Este es el origen de las familias multigénicas (Histonas, rRNAs, etc.) y de las familias génicas con un origen evolutivo común (Ej, haptoglobinas). La estructuracitogenética de las familias multigénicas suele ser muy típica: Todos los genes que componen la familia se encuentran juntos en el cromosoma en un mismo "nicho" o cluster, que a su vez puede estar repetido una o varias veces.
 |
No hay comentarios:
Publicar un comentario