Otros procesos biológicos [ editar ]
La imagen tradicional de la función de actina se relaciona con el mantenimiento del citoesqueleto y, por lo tanto, con la organización y el movimiento de los orgánulos, así como con la determinación de la forma de una célula. [79] Sin embargo, la actina tiene un papel más amplio en la fisiología de las células eucariotas, además de funciones similares en los procariotas .
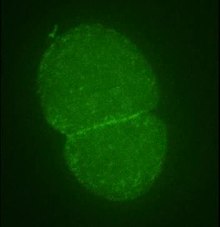
- Citocinesis . La división celular en células animales y levaduras normalmente involucra la separación de la célula madre en dos células hijas a través de la constricción de la circunferencia central. Este proceso involucra un anillo de constricción compuesto de actina, miosina y α-actinina . [131] En la levadura de fisión Schizosaccharomyces pombe , la actina se forma activamente en el anillo de constricción con la participación de Arp3 , la formina Cdc12, profilina y WASp , junto con los microfilamentos preformados. Una vez que se ha construido el anillo, la estructura se mantiene mediante un montaje y desmontaje continuo que, ayudado por Arp2 / 3Complejo y forminas, es clave para uno de los procesos centrales de la citocinesis. [132] La totalidad del anillo contráctil, el aparato del huso , los microtúbulos y el material periférico denso se denomina "cuerpo de Fleming" o "cuerpo intermedio". [79]
- La apoptosis . Durante la muerte celular programada, la familia de proteasas ICE / ced-3 (una de las proteasas convertidoras de interleuquina-1β) degradan la actina en dos fragmentos in vivo ; uno de los fragmentos es de 15 kDa y el otro de 31 kDa. Esto representa uno de los mecanismos involucrados en la destrucción de la viabilidad celular que forman la base de la apoptosis. [133] La proteasa calpaína también ha demostrado estar involucrada en este tipo de destrucción celular; [134] al igual que se ha demostrado que el uso de inhibidores de calpaína disminuye la proteolisis de la actina y la degradación del ADN (otro de los elementos característicos de la apoptosis). [135] Por otro lado, el estrés.El desencadenamiento inducido de la apoptosis causa la reorganización del citoesqueleto de actina (que también implica su polimerización), dando lugar a estructuras llamadas fibras de estrés ; Esto se activa por la vía de la quinasa MAP . [136]

- Adhesión y desarrollo celular . La adhesión entre las células es una característica de los organismos multicelulares que permite laespecialización del tejido y, por lo tanto, aumenta la complejidad celular. La adhesión de los epitelios celulares implica el citoesqueleto de actina en cada una de las células unidas, así como las cadherinas que actúan como elementos extracelulares con la conexión entre las dos mediadas por cateninas . [137] Además de la adhesión célula-célula, hay una adhesión de la célula-ECM (matriz extracelular) que se requiere para la detección de rigidez o rigidez del sustrato. Esta adhesión está mediada principalmente por la proteína adaptadora de adhesión focal llamada talina. [138]Interferir en la dinámica de la actina tiene repercusiones en el desarrollo de un organismo, de hecho, la actina es un elemento tan crucial que los sistemas de genes redundantes están disponibles. Por ejemplo, si el gen del factor de gelificación o α-actinina se eliminó en Dictyostelium, losindividuos no muestran un fenotipo anómalo, posiblemente debido al hecho de que cada una de las proteínas puede realizar la función de la otra. Sin embargo, el desarrollo de dobles mutaciones que carecen de ambos tipos de genes se ve afectado. [139]
- Modulación de la expresión génica . El estado de polimerización de Actin afecta el patrón de expresión génica. En 1997, se descubrió que la despolimerización mediada por citocalasina D en células de Schwann causa un patrón específico de expresión para los genes implicados en la mielinización de este tipo de células nerviosas . [140] Se ha demostrado que la F-actina modifica el transcriptoma en algunas de las etapas de la vida de los organismos unicelulares, como el hongo Candida albicans . [141] Además, las proteínas que son similares a la actina desempeñan un papel regulador durante la espermatogénesis en ratones [142]y, en las levaduras, se cree que las proteínas de tipo actina desempeñan un papel en la regulación de la expresión génica . [143] De hecho, la actina es capaz de actuar como un iniciador de la transcripción cuando reacciona con un tipo de miosina nuclear que interactúa con las ARN polimerasas y otras enzimas involucradas en el proceso de transcripción. [75]
- Dinámica de estereocilia . Algunas células desarrollan crecimientos filiformes finos en su superficie que tienen una función mecanosensorial . Por ejemplo, este tipo de orgánulo está presente en el órgano de Corti , que se encuentra en el oído . La principal característica de estas estructuras es que su longitud puede ser modificada. [144] La arquitectura molecular de los estereocilios incluye un núcleo de actina paracristalina en equilibrio dinámico con los monómeros presentes en el citosol adyacente. Las miosinas de tipo VI y VIIa están presentes en todo este núcleo, mientras que la miosina XVa está presente en sus extremidades en cantidades que son proporcionales a la longitud de los estereocilios. [145]
- La quiralidad intrínseca . Las redes de actomiosina se han implicado en la generación de una quiralidad intrínseca en células individuales. [146] Las células que crecen en las superficies quirales pueden mostrar un sesgo direccional hacia la izquierda / derecha que depende de la actomiosina. [147] [148]
Patología molecular [ editar ]
La mayoría de los mamíferos poseen seis genes de actina diferentes . De estos, dos codifican el citoesqueleto ( ACTB y ACTG1 ), mientras que los otros cuatro están involucrados en el músculo estriado esquelético ( ACTA1 ), el tejido muscular liso ( ACTA2 ), los músculos intestinales ( ACTG2 ) y el músculo cardíaco ( ACTC1 ). La actina en el citoesqueleto está involucrada en los mecanismos patógenos de muchos agentes infecciosos , incluido el VIH . La gran mayoría de losLas mutaciones que afectan a la actina son mutaciones puntuales que tienen un efecto dominante , con la excepción de seis mutaciones involucradas en la miopatía nemalina . Esto se debe a que, en muchos casos, el mutante del monómero de actina actúa como un "cap" al evitar el alargamiento de la actina F. [28]
Patología asociada a ACTA1 [ editar ]
ACTA1 es el gen que codifica la α- isoforma de actina que predomina en los músculos estriados esqueléticoshumanos, aunque también se expresa en el músculo cardíaco y en la glándula tiroides . [149] Su secuencia de ADN consta de siete exones que producen cinco transcripciones conocidas. [150] La mayoría de estos consiste en mutaciones puntuales que causan la sustitución de aminoácidos . Las mutaciones se asocian en muchos casos con un fenotipo que determina la gravedad y el curso de la aflicción. [28] [150]

La mutación altera la estructura y la función de los músculos esqueléticos que producen una de las tres formas de miopatía : miopatía nemalina tipo 3 , miopatía congénita con un exceso de miofilamentos delgados (CM) y miopatía congénita con desproporción del tipo de fibra(CMFT). También se han encontrado mutaciones que producen miopatías centrales . [152] Aunque sus fenotipos son similares, además de la miopatía nemalina típica, algunos especialistas distinguen otro tipo de miopatía llamada miopatía nemalina actínica. En el primero, se forman grupos de actina en lugar de las barras típicas. Es importante indicar que un paciente puede mostrar más de uno de estos fenotipos en unaLa biopsia . [153] Los síntomas más comunes consisten en una morfología facial típica (caras miopáticas), debilidad muscular, retraso en el desarrollo motor y dificultades respiratorias. El curso de la enfermedad, su gravedad y la edad en que aparece son variables, y también se encuentran formas superpuestas de miopatía. Un síntoma de la miopatía nemalínica es que las "varillas de nemalina" aparecen en diferentes lugares en las fibras musculares tipo 1. Estas barras son estructuras no patognomónicas que tienen una composición similar a la de los discos Z que se encuentran en el sarcómero . [154]
La patogenia de esta miopatía es muy variada. Muchas mutaciones ocurren en la región de la muesca de actina cerca de sus sitios de unión a nucleótidos , mientras que otras ocurren en el Dominio 2, o en las áreas donde se produce la interacción con proteínas asociadas. Esto explica de alguna manera la gran variedad de grupos que se forman en estos casos, como los cuerpos nemalinos o intranucleares o los cuerpos de cebra. [28] Los cambios en el plegamiento de la actina se producen en la miopatía nemalina, así como los cambios en su agregación y también hay cambios en la expresión de otras proteínas asociadas. En algunas variantes donde se encuentran cuerpos intranucleares, los cambios en el plegamiento enmascaran la señal de exportación de proteínas del núcleo.de modo que la acumulación de la forma mutada de actina se produce en el núcleo celular . [155] Por otra parte, parece que las mutaciones en ACTA1 que dan lugar a un CFTDM tienen un mayor efecto sobre la función sarcomérica que sobre su estructura. [156] Investigaciones recientes han tratado de comprender esta aparente paradoja, lo que sugiere que no existe una correlación clara entre el número de bastones y la debilidad muscular. Parece que algunas mutaciones son capaces de inducir una mayor tasa de apoptosis en las fibras musculares tipo II. [37]

En el músculo liso [ editar ]
ACTG2 codifica la isoforma de actina más grande, que tiene nueve exones , uno de los cuales, el que se encuentra en el extremo 5 ', no está traducido . [157] Es una γ-actina que se expresa en el músculo liso entérico. No se han encontrado mutaciones en este gen que correspondan a patologías, aunque los microarrays han demostrado que esta proteína se expresa más a menudo en casos que son resistentes a la quimioterapia con cisplatino . [158]
ACTA2 codifica una α-actina localizada en el músculo liso y también en el músculo liso vascular. Se ha observado que la mutación MYH11 podría ser responsable de al menos el 14% de los aneurismas aórticos torácicoshereditarios,particularmente el Tipo 6. Esto se debe a que la variante mutada produce un ensamblaje filamentoso incorrecto y una capacidad reducida para la contracción del músculo liso vascular. La degradación de los medios aórticos se ha registrado en estos individuos, con áreas de desorganización e hiperplasia , así como estenosisdel vasa vasorum de la aorta. [159] El número de aflicciones en las que está implicado el gen está aumentando. Se ha relacionado con la enfermedad de Moyamoya.y parece probable que ciertas mutaciones en la heterocigosis puedan conferir una predisposición a muchas patologías vasculares, como el aneurisma de aorta torácica y la cardiopatía isquémica . [160] La α-actina que se encuentra en los músculos lisos también es un marcador interesante para evaluar el progreso de la cirrosis hepática . [161]
En el músculo cardíaco [ editar ]
El gen ACTC1 codifica la isoforma α-actina presente en el músculo cardíaco. Hamada y sus compañeros de trabajo lo secuenciaron por primera vez en 1982, cuando se descubrió que estaba interrumpido por cinco intrones. [162] Fue el primero de los seis genes donde se encontraron alelos implicados en los procesos patológicos. [163]

Un número de trastornos estructurales asociados con mutaciones puntuales de este gen se han descrito que causan mal funcionamiento del corazón, tales como el tipo 1R miocardiopatía dilatada y Tipo 11 miocardiopatía hipertrófica . Ciertos defectos del tabique auricular se han descrito recientemente que también podrían estar relacionados con estas mutaciones. [165] [166]
Dos casos de cardiomiopatía dilatada se han estudiado que implica una sustitución de altamente conservadas aminoácidos pertenecientes a los dominios de la proteína que se unen y intercalan con los discos Z . Esto ha llevado a la teoría de que la dilatación se produce por un defecto en la transmisión de la fuerza contráctil en los miocitos . [30] [163]
Las mutaciones en ACTC1 son responsables de al menos el 5% de las miocardiopatías hipertróficas. [167] También se ha encontrado la existencia de una serie de mutaciones puntuales: [168]
- Mutación E101K: cambios en la carga neta y formación de un enlace electrostático débil en el sitio de unión a actomiosina.
- P166A: zona de interacción entre los monómeros de actina.
- A333P: zona de interacción actina-miosina.
La patogénesis parece implicar un mecanismo compensatorio: las proteínas mutadas actúan como toxinas con un efecto dominante, lo que disminuye la capacidad del corazón para contraerse y causa un comportamiento mecánico anormal, de modo que la hipertrofia, que generalmente se retrasa, es una consecuencia de la respuesta normal del músculo cardíaco al estrés. . [169]
Estudios recientes han descubierto mutaciones ACTC1 que están implicados en otros dos procesos patológicos: idiopática infantil cardiomiopatía restrictiva , [170] y no compactación del miocardio ventricular izquierda . [171]
En actins citoplasmáticos [ editar ]
ACTB es un muy complejo locus . Existen varios pseudogenes que están distribuidos por todo el genoma , y su secuencia contiene seis exones que pueden dar lugar a hasta 21 transcripciones diferentes mediante el empalme alternativo , que se conocen como las β-actinas. Consistente con esta complejidad, sus productos también se encuentran en varias ubicaciones y forman parte de una amplia variedad de procesos ( citoesqueleto , complejo de histona -aciltransferasa de NuA4, núcleo celular ) y, además, están asociados con los mecanismos de un gran número de procesos patológicos ( carcinomas , distonía juvenil, mecanismos de infección, malformaciones del sistema nervioso e invasión tumoral, entre otros). [172] Se descubrió una nueva forma de actina, la kappa actina, que parece sustituir al β-actina en los procesos relacionados con los tumores . [173]

Hasta ahora se han descubierto tres procesos patológicos que son causados por una alteración directa en la secuencia de genes:
- El desplazamiento de hemangiopericitoma con t (7; 12) (p22; q13) es una afección rara, en la que una mutación translocacional causa la fusión del gen ACTB sobre GLI1 en el cromosoma 12 . [175]
- La distonía de inicio juvenil es una enfermedad degenerativa rara que afecta el sistema nervioso central ; en particular, afecta áreas del neocórtex y el tálamo , donde se forman inclusiones eosinófilas en forma de varilla . Los individuos afectados representan un fenotipo con deformidades en la línea media, pérdida auditiva sensorial y distonía. Es causada por una mutación puntual en la que el aminoácido triptófanoreemplaza a la arginina en la posición 183. Esto altera la interacción de la actina con el sistema ADF / cofilina , que regula la dinámica de las células nerviosas.Formación del citoesqueleto. [176]
- También se ha descubierto una mutación puntual dominante que causa disfunción de granulocitos de neutrófilos e infecciones recurrentes . Parece que la mutación modifica el dominio responsable de la unión entre la profilina y otras proteínas reguladoras. La afinidad de Actin por la profilina se reduce considerablemente en este alelo. [177]
El locus ACTG1 codifica la proteína citosólica γ-actina que es responsable de la formación de microfilamentos del citoesqueleto . Contiene seis exones , lo que da lugar a 22 ARNm diferentes , que producen cuatro isoformascompletas cuya forma de expresión probablemente depende del tipo de tejido en el que se encuentran. También tiene dos promotores de ADN diferentes . [178] Se ha observado que las secuencias traducidas de este locus y de la β-actina son muy similares a las predichas, lo que sugiere una secuencia ancestral común que sufrió duplicación y conversión genética. [179]
En términos de patología, se ha asociado con procesos tales como amiloidosis , retinitis pigmentosa , mecanismos de infección, enfermedades renales y diversos tipos de pérdida auditiva congénita. [178]
Se ha encontrado que seis mutaciones puntuales autosómicas dominantes en la secuencia causan varios tipos de pérdida auditiva, particularmente pérdida auditiva neurosensorial relacionada con el locus DFNA 20/26. Parece que afectan a los estereocilios de las células ciliadas presentes en el órgano de Corti del oído interno . La β-actina es la proteína más abundante que se encuentra en el tejido humano, pero no es muy abundante en las células ciliadas, lo que explica la ubicación de la patología. Por otro lado, parece que la mayoría de estas mutaciones afectan las áreas involucradas en el enlace con otras proteínas, en particular la actomiosina. [28]Algunos experimentos han sugerido que el mecanismo patológico para este tipo de pérdida auditiva se relaciona con la actina F en las mutaciones que son más sensibles a la cofilina de lo normal. [180]
Sin embargo, aunque no hay registro de ningún caso, se sabe que la γ-actina también se expresa en los músculos esqueléticos, y aunque está presente en pequeñas cantidades, los organismos modelo han demostrado que su ausencia puede dar lugar a miopatías. [181]
Otros mecanismos patológicos [ editar ]
Algunos agentes infecciosos utilizan la actina, especialmente la actina citoplasmática, en su ciclo vital . Dos formas básicas están presentes en las bacterias :
- Listeria monocytogenes , algunas especies de Rickettsia , Shigella flexneri y otros gérmenes intracelulares se escapan de las vacuolas fagocíticas al recubrirse con una cápsula de filamentos de actina. L. monocytogenesy S. flexneri generan una cola en forma de "cola de cometa" que les da movilidad. Cada especie exhibe pequeñas diferencias en el mecanismo de polimerización molecular de sus "colas de cometas". Se han observado diferentes velocidades de desplazamiento, por ejemplo,encontrándose que Listeria y Shigella son las más rápidas. [182] Muchos experimentos han demostrado este mecanismo in vitro.. Esto indica que las bacterias no están usando un motor de proteína similar a la miosina, y parece que su propulsión se adquiere a partir de la presión ejercida por la polimerización que tiene lugar cerca de la pared celular del microorganismo. Las bacterias han estado rodeadas previamente por ABP del huésped y, como mínimo, el recubrimiento contiene el complejo Arp2 / 3 , las proteínas Ena / VASP , la cofilina, una proteína tamponadora y promotores de la nucleación, como el complejo de vinculina . A través de estos movimientos, forman protuberancias que llegan a las células vecinas, infectándolas también para que el sistema inmunológico solo pueda combatir la infección a través de la inmunidad celular. El movimiento podría ser causado por la modificación de la curva y el desramado de los filamentos.[183] Otras especies, como Mycobacterium marinum y Burkholderia pseudomallei , también son capaces de polimerizar localmente la actina celular para ayudar a su movimiento a través de un mecanismo centrado en el complejo Arp2 / 3. Además, el virus de la vacuna Vaccinia también utiliza elementos del citoesqueleto de actina para su difusión. [184]
- Pseudomonas aeruginosa es capaz de formar una biopelícula protectorapara escapar delas defensas deunorganismo huésped , especialmente los glóbulos blancos y los antibióticos . La biopelícula se construye utilizando ADN y filamentos de actina del organismo huésped. [185]
Además del ejemplo citado anteriormente, la polimerización de actina se estimula en los pasos iniciales de la internalización de algunos virus, en particular el VIH , por ejemplo, inactivando el complejo de cofilina. [186]
El papel que desempeña la actina en el proceso de invasión de las células cancerosas todavía no se ha determinado. [187]
Evolución [ editar ]
El citoesqueleto eucariótico de organismos entre todos los grupos taxonómicos tiene componentes similares a la actina y la tubulina. Por ejemplo, la proteína que está codificada por el gen ACTG2 en humanos es completamente equivalente a los homólogos presentes en ratas y ratones, aunque a nivel de nucleótidos , la similitud disminuye a 92%. [157] Sin embargo, hay diferencias importantes con los equivalentes en procariotas ( FtsZ y MreB ), donde la similitud entre las secuencias de nucleótidos es entre 40 y 50% entre diferentes bacteriasy arqueas.especies. Algunos autores sugieren que la proteína ancestral que dio lugar al modelo de actina eucariota se parece a las proteínas presentes en los citoesqueletos bacterianos modernos. [4] [188]

Algunos autores señalan que el comportamiento de la actina, la tubulina y la histona , una proteína involucrada en la estabilización y regulación del ADN, es similar en su capacidad para unirse a los nucleótidos y en su capacidad para aprovechar el movimiento browniano . También se ha sugerido que todos ellos tienen un antepasado común. [189] Por lo tanto, los procesos evolutivos dieron como resultado la diversificación de proteínas ancestrales en las variedades presentes en la actualidad, conservando, entre otras, las actinas como moléculas eficientes que fueron capaces de abordar procesos biológicos ancestrales esenciales, como la endocitosis . [190]
Equivalentes en bacterias [ editar ]
El citoesqueleto bacteriano puede no ser tan complejo como el que se encuentra en los eucariotas ; sin embargo, contiene proteínas que son muy similares a los monómeros y polímeros de actina. La proteína bacteriana MreB se polimeriza en finos filamentos no helicoidales y, en ocasiones, en estructuras helicoidales similares a la F-actina. [17] [191] Además, su estructura cristalina es muy similar a la de la actina G (en términos de su conformación tridimensional), incluso hay similitudes entre los protofilamentos de MreB y la actina F. El citoesqueleto bacteriano también contiene las proteínas FtsZ , que son similares a la tubulina . [192]
Las bacterias, por lo tanto, poseen un citoesqueleto con elementos homólogos a la actina (por ejemplo, MreB, ParM y MamK), aunque la secuencia de aminoácidos de estas proteínas difiere de la presente en las células animales. Sin embargo, MreB y ParM tienen un alto grado de similitud estructural con la actina eucariota. Los microfilamentos altamente dinámicos formados por la agregación de MreB y ParM son esenciales para la viabilidad celular y están involucrados en la morfogénesis celular, la segregación de cromosomas y la polaridad celular. ParM es un homólogo de actina que está codificado en un plásmido y está involucrado en la regulación del ADN plasmídico. [4] [193] Los ParMs de diferentes plásmidos bacterianos pueden formar estructuras helicoidales asombrosamente diversas que comprenden dos[194] [195] o cuatro [196] cadenas para mantener la herencia del plásmido fiel.
Aplicaciones [ editar ]
La actina se usa en laboratorios científicos y tecnológicos como una pista para motores moleculares como la miosina (ya sea en el tejido muscular o fuera de ella) y como un componente necesario para el funcionamiento celular. También se puede utilizar como herramienta de diagnóstico, ya que varias de sus variantes anómalas están relacionadas con la aparición de patologías específicas.
- Nanotecnología . Los sistemas de actina-miosina actúan como motores moleculares que permiten el transporte de vesículas y orgánulos a través del citoplasma. Es posible que la actina pueda aplicarse a la nanotecnología, ya que su capacidad dinámica se ha aprovechado en varios experimentos, incluidos los realizados en sistemas acelulares. La idea subyacente es utilizar los microfilamentos como pistas para guiar los motores moleculares que pueden transportar una carga determinada. Es decir, la actina podría usarse para definir un circuito a lo largo del cual una carga puede ser transportada de una manera más o menos controlada y dirigida. En términos de aplicaciones generales, podría utilizarse para el transporte dirigido de moléculas para depósito en lugares determinados, lo que permitiría el ensamblaje controlado de nanoestructuras. [197] Estos atributos podrían aplicarse a procesos de laboratorio como en laboratorio en un chip , en mecánica de nanocomponentes y en nanotransformadores que convierten la energía mecánica en energía eléctrica. [198]
- Control interno de las técnicas utilizadas en biología molecular , como Western blot y PCR cuantitativa . Como la actina es esencial para la supervivencia celular, se ha postulado que la cantidad de actina está bajo un control tan estricto a nivel celular que puede suponerse que su transcripción (es decir, el grado en que se expresan sus genes ) y la traducción , que Es la producción de proteínas, es prácticamente constante e independiente de las condiciones experimentales. Por lo tanto, es una práctica común en estudios de cuantificación de proteínas ( western blot ) y estudios de transcripción ( PCR cuantitativa).) para llevar a cabo la cuantificación del gen de interés y también la cuantificación de un gen de referencia como el que codifica la actina. Al dividir la cantidad del gen de interés por la del gen de la actina, es posible obtener una cantidad relativa que se pueda comparar entre diferentes experimentos, [199] siempre que la expresión de este último sea constante. Vale la pena señalar que la actina no siempre tiene la estabilidad deseada en su expresión génica . [200]
- Salud. Algunos alelos de la actina causan enfermedades; Por este motivo se han desarrollado técnicas para su detección. Además, la actina se puede usar como marcador indirecto en patología quirúrgica: es posible usar variaciones en el patrón de su distribución en el tejido como marcador de invasión en neoplasia , vasculitis y otras afecciones. [201] Además, debido a la estrecha asociación de la actina con el aparato de contracción muscular, sus niveles en el músculo esquelético disminuyen cuando estos tejidos se atrofian , por lo tanto, se puede utilizar como un marcador de este proceso fisiológico. [202]
- Tecnología alimentaria . Es posible determinar la calidad de ciertos alimentos procesados, como las salchichas , cuantificando la cantidad de actina presente en la carne constituyente. Tradicionalmente, se ha utilizado un método que se basa en la detección de 3-metilhistidina en muestras hidrolizadas de estos productos, ya que este compuesto está presente en la cadena pesada de actina y F-miosina (ambos son componentes principales del músculo). La generación de este compuesto en la carne se deriva de la metilación de los residuos de histidina presentes en ambas proteínas.
No hay comentarios:
Publicar un comentario