Defensa innata frente a patógenos infecciosos
Inmunidad innata frente a agentes infecciosos
2.1. Respuesta Natural o Innata
2.1.1. Reconocimiento inicial del microorganismo
Si un microorganismo atraviesa la barrera epitelial y comienza a crecer en los tejidos del huésped, en la mayoría de los casos, es reconocido por los macrófagos residentes en los tejidos. Los macrófagos se encuentran especialmente y en gran numero en el tejido conectivo, en asociación con el tracto gastrointestinal, en el pulmón, a lo largo de ciertos vasos en el hígado (células de Kupffer) y en el bazo. Los macrófagos son las primeras células que encuentran a los microorganismos en los tejidos 1. Estos macrófagos residentes en los tejidos ingieren, destruyen y producen quimioatractantes que activan al endotelio de los vasos locales estimulando la extravasación de neutrófilos y monocitos circulantes (reclutamiento celular). Estas células reclutadas amplifican la respuesta:
– Los neutrófilos ejecutan respuesta innata inmediata.
– Los monocitos se diferencian en macrófagos o en células dendríticas
- Los macrófagos participan en respuesta innata.
- Las células dendríticas que iniciarán la respuesta adaptativa.
2.1.2. La respuesta inflamatoria
En estas circunstancias, la interacción local de los microorganismos con los macrófagos, induce la producción de una respuesta inflamatoria que tiene como consecuencias inmediatas el reclutamiento de células y moléculas efectoras del sistema inmune innato a través de los vasos sanguíneos, que se extravasan al tejido inflamado.
Se produce la coagulación intravascular de la sangre en el lecho vascular distal y evita la diseminación inicial del microorganismo. Esta respuesta, actúa en un corto plazo de tiempo y permite el desarrollo de la respuesta inmune adaptativa, dirigida frente a antígenos específicos del microorganismo liberados en el tejido linfoide local y está dirigida frente a dianas específicas de los patógenos, habitualmente elimina la infección y protege al huésped de la reinfección por el mismo patógeno.
2.1.3. Reconocimiento de los microorganismos por células antígeno inespecíficas
Los macrófagos y neutrófilos reconocen a los micro-organismos por medio de receptores celulares para patones moleculares asociados a patógenos(PAMP).
· Distinguen entre las células del huésped y del patógeno porque se unen a moléculas típicas de los patógenos.
· No están distribuidos clonalmente.
· No tienen la especificidad de los receptores del Sistema Inmune adaptativo.
2.1.4. Patrones moleculares asociados a patógenos (PAMP)
Los ejemplos mejor conocidos son el LPS bacteriano, el peptidoglicano, ácidos lipoteicoicos, manosa, DNA bacteriano, RNA bacteriano y glicanos. Aunque estas estructuras son químicamente muy diferentes, todos los patrones moleculares asociados a patógenos tienen hechos comunes.
Primero, los patrones moleculares asociados a patógenos son producidos únicamente por patógenos microbianos y no por sus huéspedes. Por ejemplo, el LPS es sintetizado solamente por bacterias; el receptor reconoce el LPS y alerta al huésped de la presencia un organismo infectante.
Segundo, las estructuras reconocidas por el sisitema inmune innato son habitualmente esenciales para la supervivencia de los microorganismos o su patogenicidad.
Tercero, los patrones moleculares asociados a patógenos son habitualmente estructuras invariantes de clases enteras de patógenos. Por ejemplo, todas las bacterias gram negativas tienen LPS, y así el receptor de patrón de reconocimiento de LPS puede detectar la presencia de virtualmente cualquier infección bacteriana por gram negativos.

2.1.5. Características de los receptores para reconocimiento de PAMP
Los receptores del sistema inmune innato que están codificados en el genoma difieren de los receptores antigénicos en varias maneras importantes. Se expresan en muchas células efectoras del sistema inmune innato, macrófagos, células dendríticas, y células B, incluidas en el concepto de células presentadoras de antígeno. La expresión de los receptores de reconocimiento de patrones no es clonal, y tienen especificidades idénticas. Más aun, una vez que los receptores de reconocimiento de patrones identifican a un patrón molecular asociado a patógenos, la célula efectora es estimulada a desarrollar sus funciones efectoras inmediatamente, sin necesidad de proliferación ulterior. Este hecho permite la cinética rápida de la respuesta inmune innata.
2.1.6. Familias estructurales y funcionales de los receptores PAMP
Estructuralmente, los receptores de reconocimiento de patrones pertenecen a varias familias. Dominios ricos en leucina, dominios de lectinas calcio-dependientes, y dominios de receptores scavengers. Funcionalmente los receptores de reconocimiento de patrones pueden ser divididos en tres clases: secretados, endocíticos y señalizadores.

2.1.6.1. Receptores secretados
Las moléculas de reconocimiento de patrones secretadas funcionan como opsoninas al unirse a la pared de los patógenos y permitir su reconocimiento por el sistema complemento y células fagocíticas. Entre ellos estan los receptores del complemento, colectinas, pentraxin proteínas tales como la proteína amiloide del suero y la proteína C-reactiva, LPS binding protein (LBP), bactericidal permeability increasing protein (BPI) y las peptidoglycan recognition proteins (PGRS) son todos las proteínas secretadas. Una colectina muy importante es Mannose binding lectin (MBL), un PRR (Pattern Recognition Receptors) importante del sistema inmune innato que se une a una amplia gama de bacterias, virus, hongos y protozoos. MBL reconoce predominante ciertos grupos del azúcar en la superficie de microorganismos pero también fosfolípidos, ácidos nucléicos y proteínas no glicosiladas.

Son producidas junto con moléculas surfactantes por el hígado y secretadas al suero como reactantes de fase aguda. Se une a los carbohidratos de las bacterias gram positivas y negativas, hongos y a algunos virus y parásitos. Estas lectinas se asocian a 2 serin-proteasas que están relacionadas con las proteasas C1r y C1s de la vía clásica del complemento. Una vez activadas conducen a la rotura del C3 y a la activación de la C3 convertasa que resulta en la activación del complemento. Sin embargo, al contrario de C1 proteasas que requieren la unión a complejos antígeno-anticuerpo, estas proteasas asociadas a lectinas son activadas por la unión a ligandos microbianos.

2.1.6.2. Receptores endocíticos
Los receptores de reconocimiento de patrones endocíticos se localizan en la superficie de los fagocitos. Cuando reconocen un patrón molecular asociado a patógenos en una célula microbiana, inducen la fagocitosis de la bacteria y la liberación a los lisosomas, donde son destruidas. Las proteínas derivadas de los patógenos pueden ser procesadas, y los péptidos resultantes pueden ser presentados por moléculas del complejo mayor de histocompatibilidad (MCH) en la superficie de los macrófagos.
El receptor de manosa del macrófago, que es también un miembro de la familia de las lectinas calcio dependientes, es un receptor de reconocimiento de patrones. Reconoce específicamente los carbohidratos con un gran contenido de manosa que es característico de microorganismos y media su fagocitosis por los macrófagos. Otro receptor de reconocimiento de patrones endocítico, el receptor scavenger del macrófago, se une a la pared de la célula bacteriana, y juega un papel esencial en la eliminación de las bacterias de la circulación.
2.1.6.3. Receptores señalizadores
Tres familias de receptores median el reconocimiento de los microbios.
· TLR se definen por la presencia de un dominio TIR y LRR, reconocen múltiples patógenos y, por el especifico reclutamiento de los adaptadores MyD88, Mal, TRIF y TRAIM, dan lugar a la inducción de genes de respuesta inmune e inflamatoria. Las señales de esta respuesta son la activación de NF-kB y de la MAP quinasas, y, en el caso de los TLRs anti-virales, IRFS. Pro-IL-1beta y pro-IL-18 son importantes proteínas inducidas por TLR.
· NLRs tienen una estructura tripartita de dominio que consiste en una PYD o dominio CARD, una NBD y LRR. NOD1 y NOD2 detectan los subcomponentes del peptidoglicano, MDAP y MDP, respectivamente, resultando en la activación de NF-kB. NALP3 detecta ácido úrico, ARN bacteriano, toxinas bacterianas gram-positivas y ATP; NALP1b detecta el Bacillus anthracis y Ipaf detecta flagelina. NALP3 interactúa conNALP2, ASC y el Cardinal, que reclutan y habilitan la activación de la Caspasa-1 la cual actua en favor de la IL-1beta y pro-IL-18, lo que resulta en la producción de formas maduras productos bioactivos cuyos receptores actúan a través de MyD88.
· RLRs se definen por la presencia de un dominio CARD y helicasa, que detectan el ARN viral y reclutan el adaptador IPS-1 para activar IRFS, NF-kB y MAPK, lo que resulta en la inducción de las proteínas anti-virales, complementando la actividad de anti-TLR viral.

2.1.6.3.1. Toll like receptors (TLRs)
Los receptores señalizadores de la familia Toll reconocen patrones asociados a patógenos y activan la transducción de señal, activando NF-kB, que induce la expresión de una gran variedad de genes implicados en la respuesta inmune, incluyendo citoquinas inflamatorias. Recientemente se han identificados receptores de la familia Toll que pueden tener una función fundamental en la inducción de la respuesta inflamatoria e inmune.

El primer receptor de la familia Toll fue identificado en la drosófila como un componente de una vía de señalización que controla la polaridad dorsoventral de los embriones. El análisis de la secuencia de los genes Toll revela que codifica proteínas transmembrana con un gran dominio extracelular que contiene fragmentos repetidos ricos en leucina. La secuencia del dominio citoplasmático de la proteína Toll era similar al dominio citoplasmático del receptor de la interleuquina (IL) 1 de mamíferos. Además, el receptor de las IL-1 en humanos y el receptor Toll en la drosófila inducen señal de transducción que conduce a la activación de la transcripción de factores de la familia del factor nuclear kappa B (NF-kB). Miembros de esta familia tienen un papel en la inducción de la respuesta inflamatoria e inmune en los mamíferos. En los mamíferos se han identificado homólogos de los receptores Toll de la drosófila y se denominan receptores Toll-like (TLRs).
En los mamíferos se han identificado homólogos de los receptores Toll de la drosófila y se denominan receptores Toll-like (TLRs). El primer receptor Toll humano caracterizado (receptor Toll-like 4) induce la activación de la señalización para el NF-kB y la expresión de una gran variedad de citoquinas y moléculas coestimuladoras que son cruciales en la respuesta inmune adaptativa. Se conocen al menos dos miembros de la familia Toll, TLR4 y TLR2. TLR4 es el receptor para el LPS en el ratón. TLR2 es el receptor para dos principales patrones moleculares asociados a patógenos: peptidoglicano y lipoproteínas. Posteriormente se han reconocido al menos 10 TLRs, algunos, si no todos, están implicados, probablemente, en el reconocimiento de los principales patrones microbianos que desencadenan la respuesta inmune innata.

Los TLRs siempre están asociados a membrana
a) Membrana citoplasmática. Reconocen principalmente azucares, lípidos y proteínas de microorganismos patógenos.
b) Membranas de fagolisosoma o endosoma. Reconocen principalmente ácidos nucléicos y discriminan el ADN/ARN vírico del propio.

2.1.6.3.2. NOD like receptors (NLRs)
Los NLRs son proteínas citoplásmicas que pueden tener una variedad de funciones en la regulación de respuestas inflamatorias y apoptotica. Aproximadamente 20 de estas proteínas se han encontrado en el genoma mamífero e incluyen dos subfamilias importantes llamadas NODs y NALPs, CIITA, y algunas otras moléculas (e.g. IPAF y BIRC1).

La comprensión actual sugiere que algunas de estas proteínas reconozcan las moléculas endógenas o microbianas o que tensionen respuestas y que formen los oligómeros que activan caspases inflamatorias (e.g. caspase 1) desencadenando un proceso inflamatorio vía N-F-kB para inducir la producción de moléculas inflamatorias. Se conoce a la familia de NLR bajo diversos nombres, incluyendo la familia de CATERPILLER (o CLR) o de NOD-LRR.

A) NODs (NOD1 y NOD2).
Las señales NODs actúan vía N-F-kB, MAP-Kinasas. Los ligandos actualmente conocidos son:
– Las proteínas NOD1 reconoce una molécula llamada meso-DAP (meso-diaminopimelico), que es un peptidoglicano componente exclusivo de la pared de las bacterias Gram-
– Las proteínas NOD2 reconocen MDP intracelular (muramil dipéptido), que es un peptiglicano componente de la pared de las bacterias GRAM- y Gram+.

B) NALPS
NALPs contienen en el C-terminal secuencias repetidas ricas en lucinas (LRRs), que parecen actuar como dominio regulador y se pueden implicar en el reconocimiento de patógeno microbianos.

Hay 14 miembros de esta subfamilia en los seres humanos (llamados NALP1 a NALP14). Los activadores de NALP3 incluyen dipéptido muramil, ADN bacteriano, el ATP, toxinas, ARN de doble cadena, paramixovirus y los cristales de ácido úrico.
C) Otros NLRs
Otros NLRs como IPAF y NAIP5/Birc1e también pueden activar la caspase-1 en respuesta a Salmonelas y Legionella.
D) Inflamasoma.
• El inflamasoma es un complejo multiproteico (Casapasa-1, NLP3, IPAF, etc.) que promueve la maduración de las citoquinas de la inflamación de la familia de IL-1 (IL-1B, IL-18, e IL-33).
• TLRs o NOD induce síntesis de pro-IL-1 vía NF-kB.
• El inflamasoma procesa pro-IL1B vía caspasa-1y genera IL-1B activa.

• El inflamasoma es responsable de la activación de los procesos inflamatorios, y se ha demostrado que induce la piroptosis celular, un proceso de muerte programada distinto a la apoptosis.
2.1.6.3.3. Retinoic acid-inducible gene-I (RIG-I)-like receptors (RLRs)
El reconocimiento intracelular de RNA viral de cadena simple y doble es mediado por una RNA helicasa las cuáles reclutan factores vía el N-terminal del dominio para activar el programa antivirus génico. Tres helicases de este tipo se han descrito en mamíferos: RIG-I y MDA5 (reconociendo ‘ trifosfato-RNA 5 y dsRNA, respectivamente), que activan señales antivirales, y LGP2, que parece actuar como inhibidor dominante-negativo o represor.

2.1.7. Efectos del reconocimiento mediante receptores PAMP
2.1.7.1. Inflamación.
Estos cambios se inician y se modulan por citoquinas y quimioquinas producidas por Macrófagos, Células endoteliales y residentes liberados por el reconocimiento del germen. Hay migración de los leucocitos a los tejidos y participan mediadores inflamatorios como prostaglandinas, leucotrienos, PAF, activación del complemento sistema kinina y sistema de coagulación.

La unión del microorganismo con los receptores celulares de la inmunidad innata conduce a la fagocitosis y a la muerte del patógeno. La fagocitosis es un proceso activo, en el cual el patógeno unido al receptor, es inicialmente rodeado por la membrana celular y posteriormente internalizado en el fagosoma. Este se funde con los lisosomas, generando un fagolisosoma donde se libera el contenido del lisosoma y se destruye al patógeno.

Los macrófagos y neutrófilos producen una gran variedad de productos tóxicos que ayudan a matar a los microorganismos fagocitados. Los más importantes son los derivados del peroxido de hidrógeno (H2O2), el anión superóxido (O2-) y el oxido nítrico, que son tóxicos directos de las bacterias.

Los neutrófilos son células de vida corta que se mueren pronto después de que han completado la fagocitosis. Los neutrófilos muertos son el componente principal del pus que se forma en algunas infecciones, las bacterias que dan lugar a esas infecciones se denominan bacterias piógenas. Los macrófagos por otra parte, son células de vida larga y continúan generando nuevos lisosomas.
Como hemos señalado, la respuesta inflamatoria, inducida frente a la infección, depende de la producción de diversas citoquinas y quimioquinas que ocurre después del reconocimiento de los patógenos por el macrófago. Las citoquinas y quimioquinas son proteínas liberadas por distintas células después de un estimulo activador que inducen sus acciones mediante la unión a receptores específicos. Pueden actuar de forma autocrina, paracrina o endocrina. Las quimioquinas se caracterizan, además, por su actividad quimioatractiva, induciendo la emigración de aquellas células que poseen receptores apropiados.
Las citoquinas secretadas por los macrófagos en respuesta a la infección constituyen un grupo estructuralmente distinto de moléculas que incluyen, entre otras a la IL-1, 6, 10, 12, 18, TNF-alfa y las quimioquinas IL-8 y MCP-1 y que poseen numerosas actividades biológicas que favorecen el desarrollo de la respuesta inflamatoria local de carácter defensivo, pero que, como en otras circunstancias, se puede acompañar de daño tisular local y sistémico.
2.1.7.2. Efectos de la citoquinas
El TNF-alfa, la IL-1beta y otras citoquinas inducen la activación de las células endoteliales y la expresión de moléculas implicadas en la adhesión y reclutamiento leucocitario como las selectinas P y E y la molécula de adhesión VCAM-1. Además de sus efectos locales, TNF-alfa, IL-1beta e IL6, inducen rápidamente la síntesis, por los hepatocitos, de proteínas como la Proteína C Reactiva (PCR), la lectina unida a manosa y el surfactante pulmonar, denominados reactantes de fase aguda.
Estas moléculas tienen una amplia especificidad de reconocimiento de patrones asociados a patógenos y actúan, no solo como opsoninas, induciendo la fagocitosis, sino que tienen la capacidad de activar la cascada del complemento mediante la unión al fragmento C1q. Poseen, así, propiedades funcionales de los anticuerpos y se pueden unir a un gran número de microorganismos patógenos, pero no tienen la diversidad estructural de aquellos y su síntesis no es inducida de forma específica, sino en respuesta a cualquier estímulo que desencadene la liberación de TNF-alfa, IL-1 e IL-6.
Desde el punto de vista fisiopatológico, la liberación de estas tres citoquinas por los monocitos, en respuesta a la infección, participa en el desarrollo de manifestaciones clínicas de la enfermedad como la fiebre, anemia infamatoria, trombopenia, hipergammaglobulinemia y la presencia de inmunocomplejos.
El TNF-alfa, la IL-1beta y otras citoquinas inducen la activación de las células endoteliales y la expresión de moléculas implicadas en la adhesión y reclutamiento leucocitario como las selectinas P y E y la molécula de adhesión VCAM-1. Además de sus efectos locales, TNF-alfa, IL-1beta e IL6, inducen rápidamente la síntesis, por los hepatocitos, de proteínas como la Proteína C Reactiva (PCR), la lectina unida a manosa y el surfactante pulmonar, denominados reactantes de fase aguda.
Estas moléculas tienen una amplia especificidad de reconocimiento de patrones asociados a patógenos y actúan, no solo como opsoninas, induciendo la fagocitosis, sino que tienen la capacidad de activar la cascada del complemento mediante la unión al fragmento C1q. Poseen, así, propiedades funcionales de los anticuerpos y se pueden unir a un gran número de microorganismos patógenos, pero no tienen la diversidad estructural de aquellos y su síntesis no es inducida de forma específica, sino en respuesta a cualquier estímulo que desencadene la liberación de TNF-alfa, IL-1 e IL-6.
Desde el punto de vista fisiopatológico, la liberación de estas tres citoquinas por los monocitos, en respuesta a la infección, participa en el desarrollo de manifestaciones clínicas de la enfermedad como la fiebre, anemia infamatoria, trombopenia, hipergammaglobulinemia y la presencia de inmunocomplejos.
Las citoquinas liberadas por los fagocitos activan los reactantes de fase aguda.
Las bacterias inducen a los macrófagos a producir IL-6, que actúa sobre los hepatocitos para inducir la síntesis de proteínas de fase aguda (Proteína C reactiva, Proteína amiloide del suero, lectina de unión a manosa, fibrinógeno, Surfactante A y D, etc.).

Los macrófagos una vez activados, secretan un serie de citoquinas que producen efectos importantes en el organismo.

2.1.7.3. Estrategias para evadir la respuesta innata
Un hecho central que distingue a los microorganismos patógenos de los no patógenos es su capacidad para sobrepasar las defensas inmunes innatas. Los patógenos han desarrollado una gran variedad de estrategias para evitar ser inmediatamente destruidos por los macrófagos. Muchas bacterias extracelulares se recubren con una gruesa cápsula de polisacáridos que evita el reconocimiento por los receptores de los fagocitos. Otras como las micobacterias, han desarrollado mecanismos para proliferar en el interior de los fagosomas de los macrófagos mediante la inhibición de la fusión con el lisosoma. Si no poseen alguna estrategia de evasión de la inmunidad innata, los microorganismos patógenos, deben penetrar en un número suficiente en el organismo para sobrepasar la capacidad de la respuesta innata defensiva del huésped y establecer un foco de infección.
Inmunidad innata frente a bacterias extracelulares
Cuando los patógenos vencen las barreras del huésped se expone a los mecanismos de defensa de la inmunidad innata. Ya en el tejido subepitelial deberán enfrentarse al sistema del complemento, a las células fagocitarias, proteasas y anticuerpos naturales.
Muchas bacterias patógenas están capacitadas para resistir la acción de estos mecanismos. Se requiere entonces la inducción de una respuesta inflamatoria, cuyo objetivo es reclutar en el sitio de la infección elementos humorales (Anticuerpos, factores del complemento, algunas citocinas�) y celulares (neutrófilos, monocitos, NK) capaces de erradicar el foco infeccioso que se ha establecido.
Los principales mecanismos innatos frente a las bacterias extracelulares son:
|
Activación del complemento, de la fagocitosis y de la inflamación
| La respuesta inmune innata se inicia con el reconocimiento de estructuras moleculares muy conservadas y presentes en grandes grupos de microorganismos denominadas 'patrones moleculares asociados con patógenos' (PAMP). Este reconocimiento lo realizan los 'receptores de reconocimiento de patrones' (RRP) del huésped. |
Los PAMP bacterianos más conocidos son:
- Lipopolisacáridos (LPS)
- Peptidoglucano
- Ácidos lipoteicoicos
- Mananos (manosa
- DNA bacteriano
- RNA de doble cadena
- Glucanos
Estos PAMP son esenciales para la supervivencia y patogenicidad de las bacterias.
Cuando los RRP reconocen un PAMP los componentes de la inmunidad innata se activan inmediatamente.
Funcionalmente, los RRP pueden dividirse en tres clases:
|
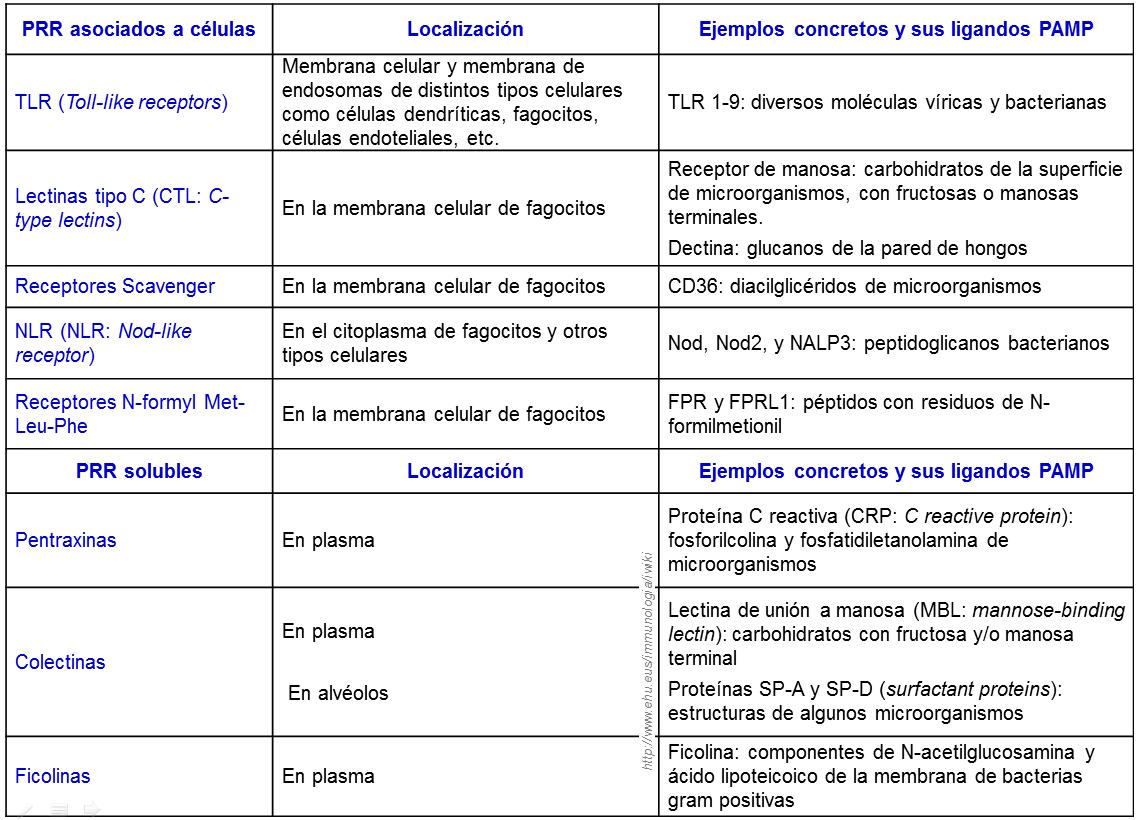
Ejemplos de moléculas de reconocimiento de la Inmunidad innata y patrones moleculares que reconocen en los microorganismos.
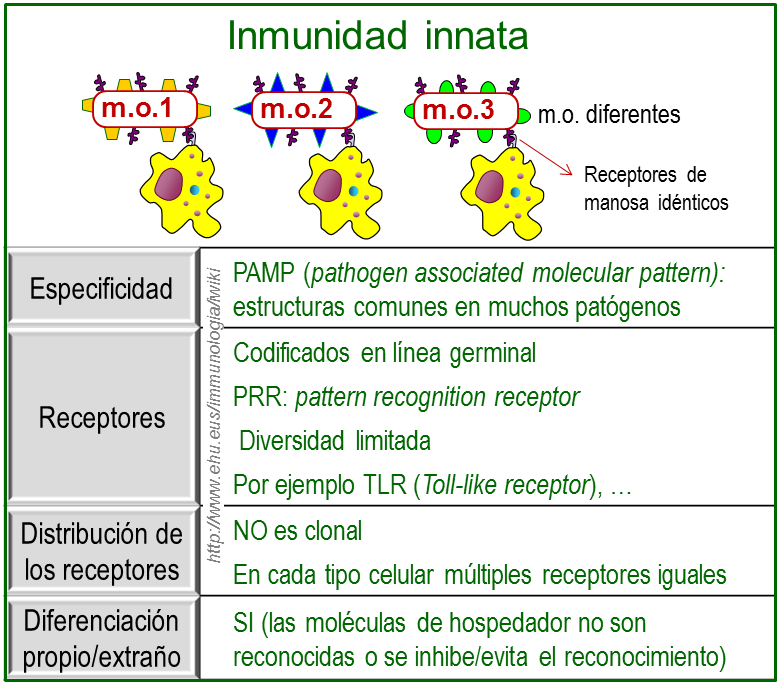
Especificidad en el reconocimiento de la inmunidad innata.
Los RRP secretados son moléculas solubles que se unen a las paredes celulares de las bacterias y funcionan como opsoninas.
Un ejemplo es la lectina de unión a manosa. Esta lectina se une a la manosa de las paredes de los microorganismos y activa el complemento por la vía de las lectinas.
Otros RRP secretados, al actuar como opsoninas, favorecen la fagocitosis del patógeno.
Los RRP endocíticos se expresan en la membrana de los fagocitos y de las células dendríticas. Al reconocer los PAMP, estos receptores activan la ingestión y degradación del patógeno en los fagolisosomas de estas células. Tras lo que los péptidos derivados del procesamiento del patógeno por estas CPA profesionales, se presentan en las moléculas de clase I y clase II (presentación cruzada) de macrófagos y células dendríticas.
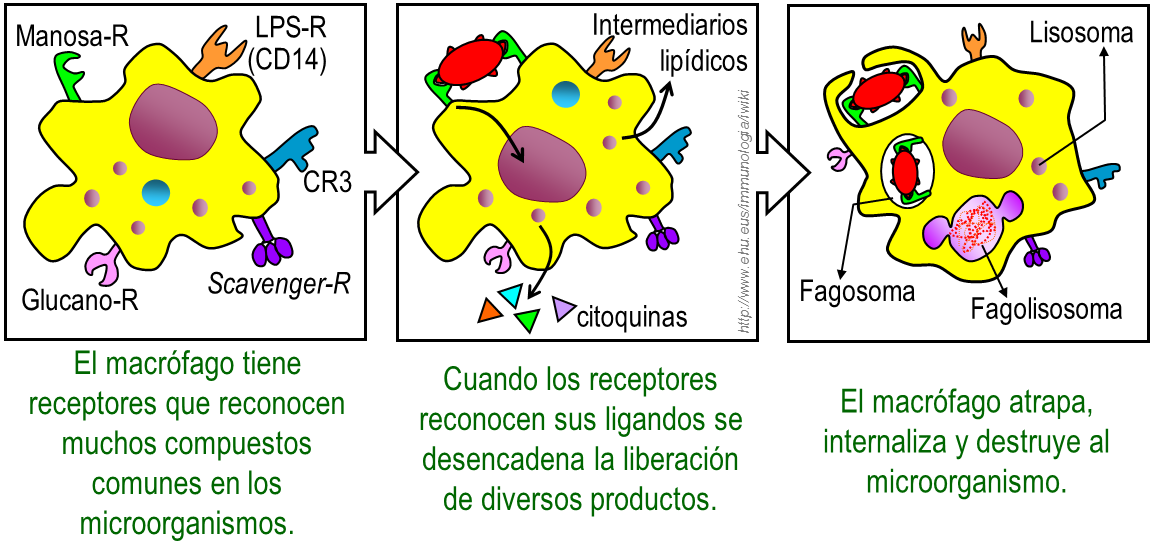 Fagocitosis mediada por fagocitos en la inmunidad innata..
Fagocitosis mediada por fagocitos en la inmunidad innata..
Los RRP de señalización (en este grupo están los receptores de la familia Toll), al reconocer a los PAMP, activan la producción de citocinas por las células de la respuesta innata. Algunas de estas citocinas son proinflamatorias (Fig1- 4).
La activación del complemento tiene un poderoso efecto antibacteriano: :
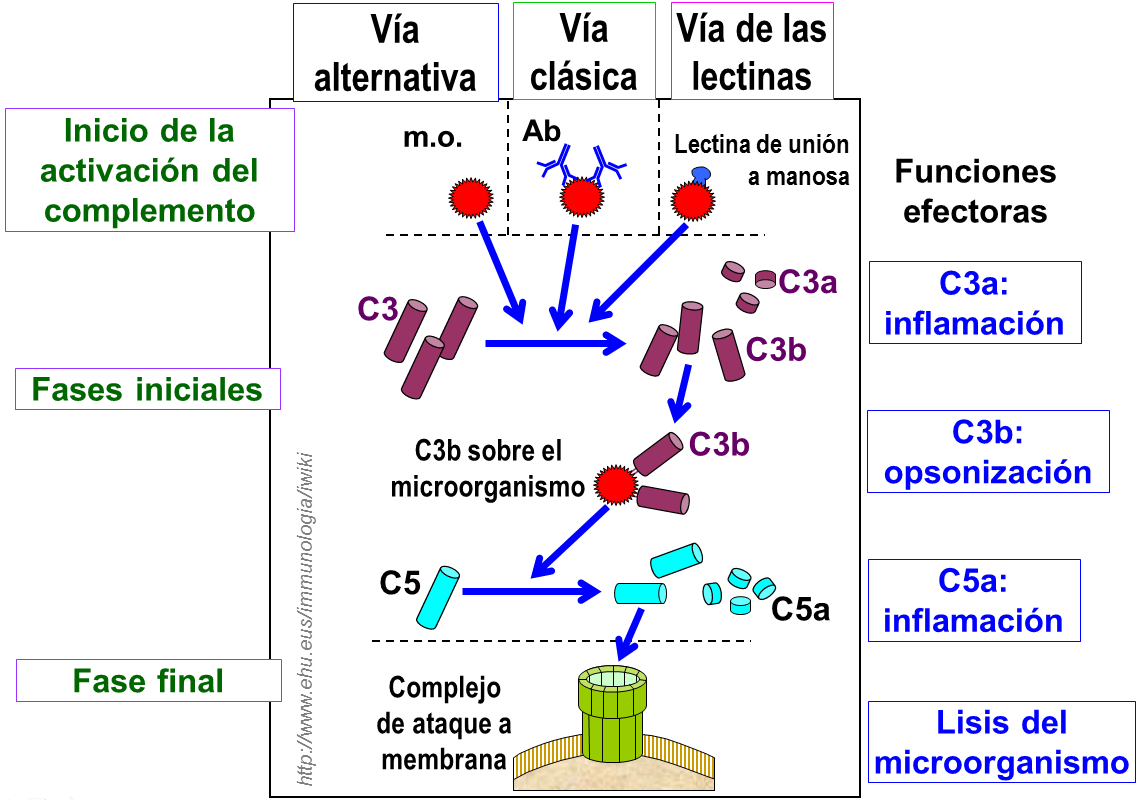
Principales actividades biológicas de las moléculas que se forman durante la activación del complemento.
- la producción de los factores C3a, C4a, y C5a median la respuesta inflamatoria, al reclutar y activar diferentes tipos celulares: neutrófilos, monocitos, macrófagos, NK, células dendríticas, eosinófilos, mastocitos, células endoteliales y células musculares lisas. El desarrollo de una respuesta inflamatoria permite reclutar componentes humorales y celulares en el foco de la infección que tratarán de eliminarla.
- la opsonización de las bacterias por C3b potencia su fagocitosis por neutrófilos y macrófagos, estas células expresan un conjunto de receptores para reconocer esta opsonina inespecífica.
- la formación del complejo de ataque a membrana lisa las bacterias gram-nagativas y entre estas especialmente a las especies de Neisseria y a Haemophilus influenzae.
- eliminación de inmunocomplejos circulantes en el hígado y el bazo mediada por el factor C3b y su unión a sus receptores en la membrana del eritrocito.
- los productos de degradación de C3b potencian la activación del linfocito B.
| El LPS de las bacterias gramnegativas y los ácidos teicoicos de las grampositivas activan el complemento por la vía alternativa |
Producción de citocinas en la respuesta innata
Las células fagocitarias presentes en el foco infeccioso se activan al reconocer los PAMP o citocinas producidas por ellas mismas (efecto autocrino) o por otros tipos celulares (efecto paracrino).
| Después del reconocimiento de la bacteria, opsonizada o no, las células fagocitarias, neutrófilos y macrófagos internalizan el patógeno para eliminarlo. Los neutrófilos son las principales células fagocitarias y los macrófagos, tras activarse al ingerir el patógeno, producen gran número de citocinas. |

Para hacer frente a ala infección, las citoquinas secretadas por los macrófagos participan en numerosas actividades biológicas y facilitan la coordinación de la respuesta frente a la infección.
Como consecuencia de su activación los macrófagos producen diferentes familias de citocinas:
- Citocinas que inducen una respuesta inflamatoria aguda, local o sistémica: IL-1 (IL-1α, IL-1β), TNF-α e IL-6.
- Citocinas que median el reclutamiento de leucocitos en el tejido inflamado: quimiocinas.
- Citocinas que inducen la proliferación y/o diferenciación de precursores leucocitarios en la médula ósea: G-CSF, M-CSF y GM-CSF.
- Citocinas que orientan el tipo de la respuesta inmune adaptativa: IL-12 e IL-18, IL-1β, TGF-β.
- Citocinas que inhiben la activación del macrófago en el foco inflamatorio, con el fin de regular estos eventos y evitar lesiones inflamatorias en el huésped: IL-10 y TGF-β
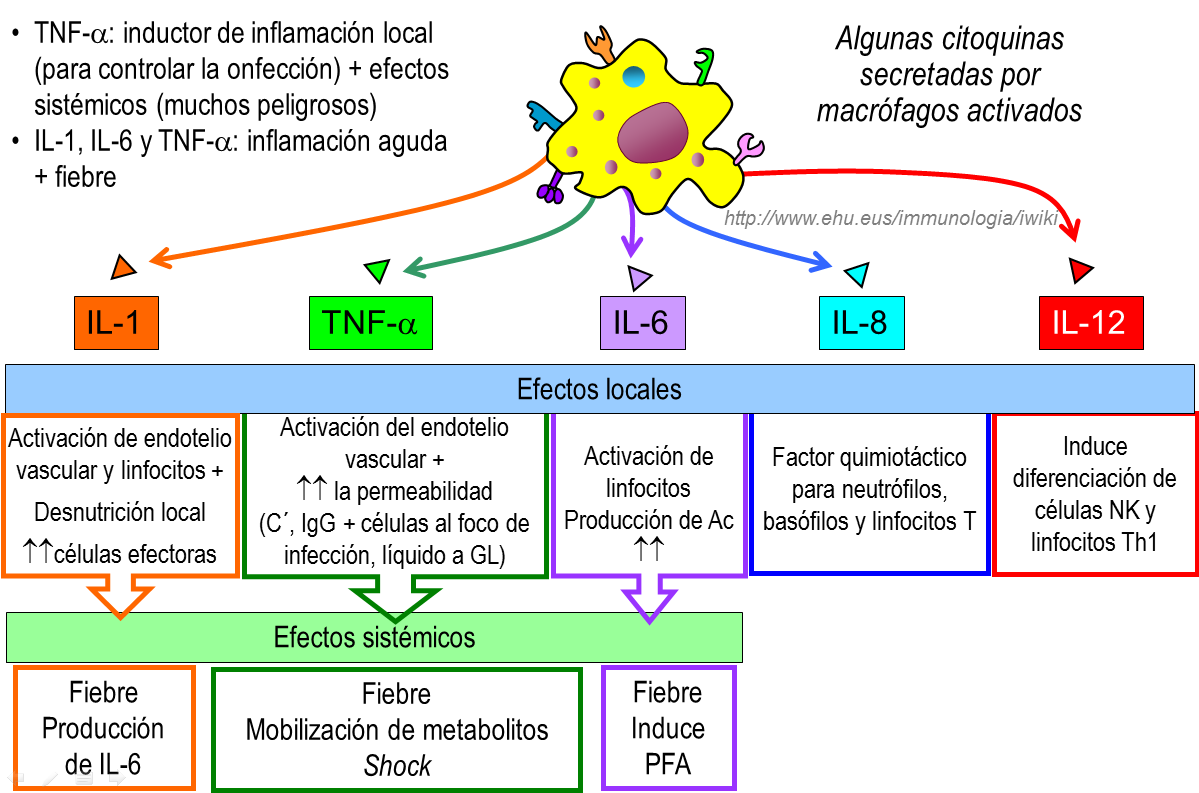
Acción de algunas de las citocinas secretadas por el macrófago para hacer frente a la infocción.
Makrofagoek ekoizten dituzten zitokinek, funtzio biologiko anitz dituzte eta infekzioen aurkako erantzunak koordinatzeko baliagarriak dira. Adibidez, IL-1, IL-6 eta TNFαzitokinak hurrengoetan eragina dute:
- Gibelean: Fase akutuko proteinen sintesia (C proteina erreaktiboa, manosa lotzen duen proteina) → Konplementua aktibatu, opsonizazioa.
- Hezur-muineko endotelioan: Neutrofiloen mobilizazioa → Fagozitosia
- Hipotalamoan: Gorputzeko tenperaturaren gehitzea → Birusen eta bakterioen erreplikazioa gutxitu; antigenoaren prozesamendua areagotu; erantzun espezifikoari lagundu.
- Koipean, muskuluan: Proteinen eta energiaren mobilizazioa gorputzeko tenperatura igotzeko → Birusen eta bakterioen erreplikazioa gutxitu; antigenoaren prozesamendua areagotu; erantzun espezifikoari lagundu.
- Zelula dendritikoetan: TNFak zelula horien heltzea eta migrazioa linfa-gongoiletara eragiten du → erantzun espezifikoa hasi.
| Durante la infección bacteriana no solo los leucocitos contribuyen a la producción de citocinas y quimiocinas, sino también las células que integran los diferentes tejidos. |
Las citocinas IL-12 y TNF-α liberadas por los macrófagos activan a las células NK, quienes responden produciendo IFN-γ TNF-α, TNF-β, IL-10, IL-13 y GM-CSF. En la defensa frente a bacterias extracelulares las NK juegan un papel importante por esta producción de citocinas: El IFN-γ que producen mantiene la activación del macrófago.

NK zelulen mekanismoak immunitate innatoan
Las células NK forman parte de la inmunidad innata y participan en la formación de una primera línea de defensa contra los patógenos mediante dos mecanismos:
|
No hay comentarios:
Publicar un comentario