apomorfia o apomorfía1 ("forma separada") es un rasgo evolutivamente novedoso, es decir, derivado de otro rasgo perteneciente a un taxón ancestral filogenéticamente próximo. El concepto de apomorfia se define en relación al deplesiomorfia, referido a aquellos rasgos históricamente antiguos, derivados, por tanto, de un taxón ancestral o primitivo.
La distinción entre apomorfia y plesiomorfia no es absoluta, sino relativa a un nivel determinado de la jerarquía sistemática:
- En el contexto de los mamíferos como subtaxón de los tetrápodos, el martillo del oído es una apomorfia. En relación a este último, el cartílago articular de los anfibios (una parte de lamandíbula inferior de la que se deriva el martillo) es un carácter plesiomórfico.
- Si consideramos a los animales como un conjunto, la columna vertebral es una apomorfia que caracteriza al subtaxón de los vertebrados. Pero si queremos caracterizar la posesión de columna vertebral en un pájaro en relación al taxón de los vertebrados, entonces el rasgo es plesiomórfico.
Cuando se somete a análisis un dendrograma, cladograma ofilograma, las apomorfías observadas (caracteres que difieren del ancestral) se pueden subdividir en dos subgrupos:
- Sinapomorfías: Aquellas apomorfías compartidas por dos o más grupos.
- Autapomorfías: Aquellas apomorfías que sólo aparecen en un grupo.

{kind=link}
Apomorfía: en cladística, estado novedoso o derivado de un carácter. Cuando es compartido por varios taxones se denomina sinapomorfía. Véase: plesiomorfía y autapomorfía.
1. Simplesiomorfía, 2. Sinapomorfía, 3. Autapomorfía. A-E: Taxones existentes, F-I: Taxones antepasados.





aptitud, adecuación biológica o eficacia biológica (en inglés fitness, denotada a menudo como 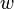 en los modelos de genética de poblaciones) es un concepto primordial en la teoría de la evolución.
en los modelos de genética de poblaciones) es un concepto primordial en la teoría de la evolución.
en los modelos de genética de poblaciones) es un concepto primordial en la teoría de la evolución.
Describe la capacidad de un individuo de reproducirse con cierto genotipo, y normalmente es igual a la proporción de los genes del individuo en los genes totales de la siguiente generación. Si las diferencias entre genotipos distintos afectan a la aptitud, entonces las frecuencias de los genotipos cambiarán a lo largo de las generaciones; los genotipos con mayor aptitud se harán más comunes. Este proceso se llama selección natural.
En términos genéticos puede definirse como la contribución promedio de un alelo a las siguientes generaciones:
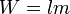
donde W=Adecuación absoluta; l=Supervivencia y m=fecundidad.
La adecuación relativa o eficacia biológica relativa es una medida utilizada al comparar varias adecuaciones donde éstas son relativizadas con respecto a alguna otra adecuación (generalmente la más alta).
Principios básicos de la genética de poblaciones
1. La genética de poblaciones es la síntesis entre la teoría darwiniana y los principios de la genética mendeliana. Los principales conceptos que estructuran los modelos de esta disciplina son los de población, reservorio génico, frecuencias genotípicas, frecuencias génicas y panmixia.
2. La población es una unidad definida por su reservorio génico, que es el conjunto de todos los alelos de todos los genes de sus individuos que la integran. Cada individuo es el depositario temporal de una pequeña muestra del reservorio génico.
3. Las características del reservorio génico pueden ser descritas a través de parámetros estadísticos que indican la frecuencia de los diferentes alelos y de sus combinaciones genotípicas. Si una población está constituida por un númeroN de organismos diploides, un número d de organismos homocigóticos dominantes, un número h de heterocigóticos y un número r de homocigóticos recesivos, entonces d + h + r = N. Las frecuencias relativas de cada genotipoestán representadas por las letras D, H y R, respectivamente.
4. Cada gameto contiene sólo uno de los dos alelos de cada uno de los sistemas genéticos que constituyen el genotipo del individuo que lo produce. En consecuencia, son los alelos y no los genotipos los que tienen continuidad hereditaria.
5. Los N individuos de una población se forman por la unión de dos copias de cada uno de los alelos, de modo que el número total de alelos en dicha población es 2N. Si cada individuo Aa tiene un solo alelo A y los homocigotas AA tienen dos alelos A, el número total de alelos A en la población es 2d + h. Si p es la frecuencia relativa de alelos A y q la proporción de alelos a, entonces p + q = 1.
6. La panmixia es un patrón reproductivo en el que los individuos se aparean al azar, con exclusión de toda preferencia fenotípica. En general, en una población, esta condición sólo se cumple para algunos genes.
Un estado estacionario: el equilibrio de Hardy-Weinberg
7. En 1908, Godfrey Hardy y Wilhelm Weinberg demostraron que, en ciertas condiciones, las combinaciones que resultan del apareamiento y reproducción en los organismos diploides no involucran un cambio en la composición general del reservorio génico de una población.
8. Las condiciones del modelo de Hardy-Weinberg son las siguientes:
- No ocurren mutaciones.
- No hay inmigración ni emigración.
- La población es lo suficientemente grande como para que se apliquen las leyes de la probabilidad.
- El apareamiento entre individuos es al azar.
- Todos los genotipos presentan el mismo éxito.
9. En una población donde se cumplen todas las condiciones del modelo, las frecuencias de los alelos A y a no cambian de una generación a otra. Tampoco cambian las frecuencias de los genotipos AA, Aa y aa.
10. El equilibrio alcanzado se expresa mediante la ecuación: p2 + 2pq + q2 = 1, donde p2 representa la frecuencia de individuos homocigóticos para un alelo, q2la frecuencia de los homocigóticos para el otro alelo y 2pq la frecuencia de los heterocigóticos. El modelo de Hardy-Weinberg se aplica también en los casos de alelos múltiples, pero la ecuación que representa el equilibrio es más compleja.
Fig. 18-2. Frecuencias de genotipos según la ley de Hardy-Weinberg

Supongamos que las frecuencias relativas de A(0,8) y a (0,2) son iguales tanto en los machos como en las hembras (como ocurre con la mayoría de los alelos en las poblaciones naturales). Supongamos ahora que los machos y las hembras se aparean al azar con respecto al hecho de ser portadores de los alelos A y a. Podemos calcular las frecuencias de los genotipos resultantes dibujando un tablero de Punnett. Las proporciones de los genotipos en la población producida por este apareamiento al azar serán 64% AA, 32% Aay 4% aa. En este caso, estamos suponiendo que el gen no está ligado al sexo y que los alelos tienen las mismas frecuencias en los machos y en las hembras.
El concepto de aptitud
11. Para la genética de poblaciones, la evolución es el cambio intergeneracional de frecuencias alélicas en las poblaciones. La aptitud (fitness) representa el éxito diferencial de los genotipos, y se mide por el número relativo de descendientes vivos que deja un individuo, que indica en qué medida sus genes estarán presentes en la siguiente generación.
La variabilidad: materia prima del cambio evolutivo
12. Las variaciones de los componentes bióticos y abióticos del ambiente ejercen presiones sobre el reservorio génico de las poblaciones.
13. En la década de 1960, Richard C. Lewontin y J. L. Hubby estudiaron la variación de enzimas individuales mediante la técnica de electroforesis. Este método subestima la variación, porque muchos cambios en la secuencia de nucleótidos no se traducen en cambios en la secuencia de la proteína o afectan regiones de DNA no codificante.
14. En la década de 1980, mediante la técnica de PCR seguida por la secuenciación del DNA amplificado, se puso de manifiesto gran cantidad de variabilidad que previamente no se había detectado. Estos estudios revelaron que las poblaciones naturales son más variables de lo que inicialmente se había estimado.
15. Dos escuelas intentan explicar los niveles elevados de polimorfismoobservados en las poblaciones. La escuela seleccionista afirma que toda variación con base genética afecta directa o indirectamente la aptitud. Los neutralistas, en cambio, sostienen que la mayor parte de las variantes genéticas son neutras o confieren diferencias tan pequeñas en la aptitud que su pérdida o preservación no depende de la selección natural. Para esta escuela, la variación intrapoblacional y la divergencia entre poblaciones son el resultado del surgimiento por mutación de nuevas variantes neutras y su pérdida a causa de la deriva génica.
Origen y preservación de la variabilidad genética
16. Las mutaciones constituyen la materia prima del cambio evolutivo, ya que proporcionan las variaciones sobre las que operan otras fuerzas evolutivas.
17. Se ha propuesto que los genes que hoy componen los genomas se originaron a partir de unos pocos protogenes que se duplicaron y se modificaron por acumulación de mutaciones. Este proceso de duplicación y subsiguiente mutación continuaría operando en el presente y podría explicar el origen y la diversificación de las diversas familias génicas que componen los genomas eucariontes.
18. La reproducción sexual, principal mecanismo que promueve la variabilidad, produce nuevas combinaciones genéticas mediante tres mecanismos:
- La distribución independiente de los cromosomas durante la meiosis.
- El entrecruzamiento.
- La combinación al azar de los genomas parentales en la fecundación.
19. La recombinación no es efectiva en presencia de homogeneidad genética, pero las poblaciones poseen mecanismos que favorecen la exogamia. Se trata de patrones de apareamiento que minimizan la probabilidad de que se apareen dos individuos emparentados.
20. La diploidia contribuye a preservar la variabilidad en los eucariontes, a través del almacenamiento de alelos recesivos en los individuos heterocigóticos. Aun tratándose de alelos que confieran desventajas a los homocigóticos, cuanto menor sea su frecuencia, menos expuestos estarán y más lentamente serán eliminados por la selección natural.
No hay comentarios:
Publicar un comentario