| Célula de Golgi | |
|---|---|

Microcircuitos del cerebelo. Las sinapsis excitatorias se indican con (+) y las sinapsis inhibitorias con (-).
MF: Fibra de musgo . DCN: Núcleos cerebelosos profundos . IO: Olivo inferior . CF: Fibra trepadora . GC: Gránulo celular . PF: fibra paralela . PC: célula de purkinje . GgC: célula de Golgi. SC: Celda estrellada . BC: cesta celular . | |
| Detalles | |
| Ubicación | Capa granular delcerebelo |
| Identificadores | |
| IdentificaciónNeuroLex | nifext_129 |
| Términos anatómicos de la neuroanatomía. | |
En neurociencia , las células de Golgi son interneuronas inhibitorias que se encuentran dentro de la capa granular del cerebelo . Fueron identificados por primera vez como inhibidores por Eccles et al. en 1964. [1] También fue el primer ejemplo de una red de retroalimentación inhibitoria, donde la interneurona inhibitoria se identificó anatómicamente. Estas células hacen sinapsis con la dendrita de células granulares y células de cepillo unipolares. Reciben información excitatoria de fibras musgosas , también sinapsis en células granulares y fibras paralelas., que son axones de células granulares largas. De este modo, esta circuitería permite la inhibición de realimentación y realimentación de las células granulares.
La sinapsis principal hecha por estas células es una sinapsis en la fibra musgo - sinapsis excitatoria de células granulares en un glomérulo. El glomérulo está formado por el terminal de fibras musgosas, las dendritas de células granulares , el terminal de Golgi y está encerrado por una capa glial. [2] La célula de Golgi actúa alterando la fibra musgosa - sinapsis de células granulares.
Las células de Golgi usan GABA como su transmisor. El nivel basal de GABA produce una conductancia de fuga postsináptica mediante la activación tónica de los receptores GABA-A que contienen alfa 6 en la célula del gránulo. [3] [4] [5] Estos receptores de alta afinidad se ubican tanto sinápticamente como extrainápticamente en la célula del gránulo. Los receptores sinápticos median la contracción fásica, la duración de alrededor de 20-30 ms, mientras que los receptores extrasinapáticos median la inhibición tónica de alrededor de 200 ms, y se activan por desbordamiento de sinapsis. [6]
Además, el GABA actúa sobre los receptores GABA-B que se ubican de forma presináptica en el terminal de fibra de musgo. Estos inhiben las EPSC evocadas de la fibra de musgo de la célula granular de manera dependiente de la temperatura y la frecuencia. A alta frecuencia de cocción de musgo (10 Hz), el GABA no actúa sobre los receptores presinápticos GABA-B en las EPSC evocadas. Sin embargo, a baja (1 Hz), el GABA tiene un efecto en las EPSC mediadas a través de estos receptores presinápticos GABA-B.
Las células de Martinotti son pequeñas neuronas multipolares con dendritas de ramificación corta . Están dispersos a lo largo de varias capas de la corteza cerebral, enviando sus axones hasta la capa cortical I, donde forman una arborización axonal . Los cenadores transgreden múltiples columnas en la capa VI y hacen contactos con las dendritas de mechones distales de las células piramidales . [1] Las células de Martinotti expresan somatostatina y algunas veces calbindina , pero no parvalbúmina o péptido intestinal vasoactivo . [2]Además, se ha demostrado que las células de Martinotti en la capa V expresan la subunidad α2 del receptor nicotínico de acetilcolina (Chrna2). [3]
Investigaciones recientes sugieren que las células de Martinotti están asociadas con un mecanismo de amortiguación cortical. Cuando la neurona piramidal , que es el tipo más común de neurona en el cerebro, comienza a sobreexcitarse, las células de Martinotti comienzan a enviar señales inhibitorias a las neuronas circundantes. [4]
Históricamente, el descubrimiento de las células de Martinotti se ha atribuido erróneamente a Giovanni Martinotti en 1888, aunque ahora se acepta que en realidad fueron descubiertos en 1889 por Carlo Martinotti (1859–1908), un estudiante de Camillo Golgi .
Las células de rueda de carro son neuronas del núcleo coclear dorsal (DCN) donde superan en gran medida a las otras interneuronas inhibitorias de la DCN. Sus somas se encuentran en el lado superficial de la capa piramidal de la DCN, y sus dendritas reciben información de las fibras paralelas de la capa de células granulares. Sus axones no se extienden más allá del núcleo coclear dorsal, sino que hacen sinapsis con otras células de rueda de carro y células piramidales dentro de la DCN que liberan GABA y glicina en sus objetivos.
Las células de rueda de carro tienen patrones de picos similares a las células de Purkinje , disparando explosiones de picos complejas, así como picos simples. También se ve que comparten otras características comunes a las células cerebrales de Purkinje. [1] Otros datos apoyan la similitud estructural y funcional encontrada entre la rueda de carro y la neurona de Purkinje. [2] Las células de la rueda de carro tienen una alta densidad de espinas en sus dendritas, sus axones son mielinizados, tienen forma de células esféricas y de tamaño mediano. [3]Los principales objetivos de las células de rueda de carro son las neuronas piramidales eferentes grandes de la DCN, que se encuentran en la capa dos. Los datos recuperados han demostrado que esta orientación de las células piramidales modula su actividad y, por lo tanto, desempeña un papel clave en la configuración de la salida global de las capas superficiales del núcleo coclear dorsal.
Las células periglomerulares median la inhibición lateral en el sistema olfativo junto con las células granulares . Tienen sinapsis inhibitorias en células mitral y células en penachos .
Las células de Purkinje , o neuronas de Purkinje , son una clase de neuronas GABAérgicas ubicadas en el cerebelo . Su nombre proviene de su descubridor, el anatomista checo Jan Evangelista Purkyně , quien caracterizó las células en 1839.
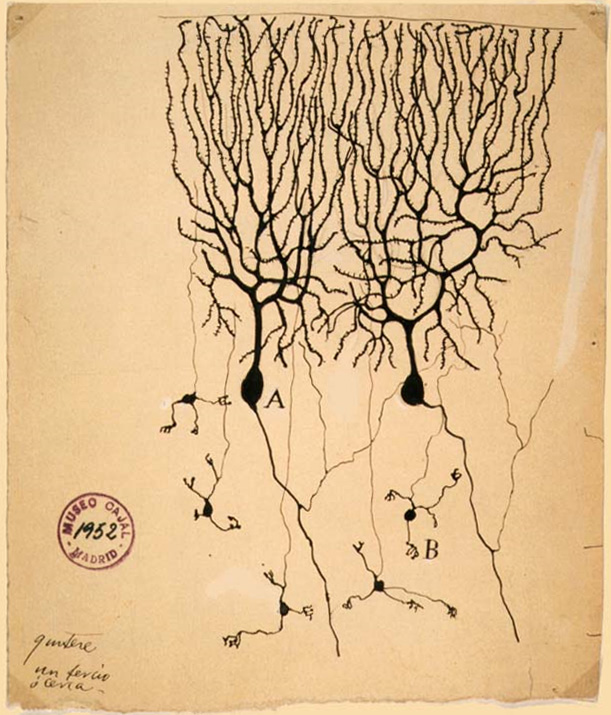
Estructura [ editar ]




Estas células son algunas de las neuronas más grandes del cerebrohumano ( las células de Betz son las más grandes), [1] con un eje dendríticointrincadamente elaborado , caracterizado por una gran cantidad de espinas dendríticas . Las células de Purkinje se encuentran dentro de la capa de Purkinje en el cerebelo . Las celdas de Purkinje están alineadas como dominós apilados uno frente al otro. Sus grandes cenizas dendríticas forman capas casi bidimensionales a través de las cuales pasan las fibras paralelas desde las capas más profundas. Estas fibras paralelas hacen relativamente más débil excitador.Las sinapsis ( glutamatérgicas ) a las espinas en la dendrita de células de Purkinje, mientras que las fibras trepadoras que se originan en el núcleo olivar inferior en la médula proporcionan una entrada excitadora muy poderosa a las dendritas proximales y al soma celular. Las fibras paralelas pasan ortogonalmente a través del eje dendrítico de la neurona de Purkinje, con hasta 200,000 fibras paralelas [2] que forman una sinapsis de células de Puruleje-células de Granule con una única célula de Purkinje. Cada célula de Purkinje recibe aproximadamente 500 sinapsis de fibras trepadoras, todas originadas de una sola fibra trepadora. [3] Células de cesta y estrelladas (encontradas en la capa molecular cerebelosa) proporcionan entrada inhibitoria (GABAergic) a la célula de Purkinje, con células de cesta sincronizadas en el segmento inicial del axón de la célula de Purkinje y células estrelladas en las dendritas.
Las células de Purkinje envían proyecciones inhibitorias a los núcleos cerebelosos profundos, y constituyen el único resultado de toda coordinación motora en la corteza del cerebelo.
Molecular [ editar ]
La capa de Purkinje del cerebelo, que contiene los cuerpos celulares de las células de Purkinje y la glía de Bergmann , expresa una gran cantidad de genes únicos. [4] También se propusieron marcadores genéticos específicos de Purkinje comparando el transcriptoma de ratones deficientes de Purkinje con el de ratones de tipo salvaje. [5] Un ejemplo ilustrativo es la proteína celular de Purkinje 4 ( PCP4 ) en ratones knockout , que muestran un aprendizaje locomotor deficiente y una plasticidad sináptica notablemente alterada en las neuronas de Purkinje. [6] [7] PCP4 acelera tanto la asociación como la disociación del calcio (Ca 2+ ) concalmodulina (CaM) en el citoplasma de las células de Purkinje, y su ausencia perjudica la fisiología de estas neuronas. [6] [7] [8] [9]
Desarrollo [ editar ]
Existe evidencia en ratones y humanos de que las células de la médula ósea se fusionan o generan células de Purkinje cerebelosas, y es posible que las células de la médula ósea, ya sea por generación directa o por fusión celular, puedan desempeñar un papel en la reparación del daño al sistema nervioso central. [10] [11] [12] [13] [14] Además la evidencia apunta todavía hacia la posibilidad de un común de células madre ancestro entre las neuronas de Purkinje, linfocitos B y aldosterona -producir células del humano corteza suprarrenal . [13]
Función [ editar ]

Las células de Purkinje muestran dos formas distintas de actividad electrofisiológica:
- Los picos simples se producen a velocidades de 17 - 150 Hz (Raman y Bean, 1999), ya sea espontáneamente o cuando las células de Purkinje se activan sinápticamente por las fibras paralelas, los axones de las células granulares.
- Los picos complejos son lentos, picos de 1–3 Hz, caracterizados por un pico inicial de gran amplitud prolongada, seguido por una explosión de alta frecuencia de potenciales de acción de menor amplitud. Son causados por la activación de la fibra trepadora y pueden implicar la generación de potenciales de acción mediados por el calcio en las dendritas. Después de la actividad compleja de picos, los picos simples pueden ser suprimidos por la poderosa entrada de picos complejos. [15]
Las células de Purkinje muestran una actividad electrofisiológica espontánea en forma de trenes de picos que dependen tanto del sodio como del calcio. Esto fue mostrado inicialmente por Rodolfo Llinas (Llinas y Hess (1977) y Llinas y Sugimori (1980)). Los canales de calcio tipo P fueron nombrados después de las células de Purkinje, donde se encontraron inicialmente (Llinas et al. 1989), que son cruciales en la función cerebelosa. Ahora sabemos que la activación de la célula de Purkinje por las fibras trepadoras puede cambiar su actividad de un estado tranquilo a un estado espontáneamente activo y viceversa, que sirve como una especie de interruptor de palanca. [dieciséis]Estos hallazgos han sido cuestionados por un estudio que sugiere que la alternancia de este tipo mediante la entrada de fibras trepadoras ocurre predominantemente en animales anestesiados y que las células de Purkinje en animales de comportamiento despierto, en general, operan casi continuamente en el norte del estado. [17] Pero este último estudio ha sido desafiado [18] y desde entonces se ha observado el cambio de células de Purkinje en gatos despiertos. [19] Un modelo computacional de la célula de Purkinje ha demostrado que los cálculos de calcio intracelular son responsables de la conmutación. [20]
Los hallazgos han sugerido que las dendritas de células de Purkinje liberan endocannabinoides que pueden regular a la baja tanto las sinapsis excitadoras como las inhibitorias. [21] El modo de actividad intrínseca de las células de Purkinje se establece y controla mediante la bomba de sodio-potasio . [22] Esto sugiere que la bomba podría no ser simplemente una molécula homeostática "de mantenimiento" para gradientes iónicos. En su lugar, podría ser un elemento de cálculo en el cerebelo y el cerebro. [23] De hecho, una mutación en el Na +.
- K +
la bomba causa la aparición rápida de parkinsonismo por distonía; Sus síntomas indican que es una patología de la computación del cerebelo. [24] Además, usando el veneno ouabain para bloquear Na +
- K +
Las bombas en el cerebelo de un ratón vivo inducen ataxia y distonía . [25] El modelado numérico de datos experimentales sugiere que, in vivo, el Na +
- K +
la bomba produce largas puntuaciones quiescentes (>> 1 s) a la activación de las neuronas de Purkinje; estos pueden tener un papel computacional. [26] El alcohol inhibe Na +
- K +
Bombea en el cerebelo y es probable que corrompa la computación del cerebelo y la coordinación corporal. [27] [28]
- K +
la bomba causa la aparición rápida de parkinsonismo por distonía; Sus síntomas indican que es una patología de la computación del cerebelo. [24] Además, usando el veneno ouabain para bloquear Na +
- K +
Las bombas en el cerebelo de un ratón vivo inducen ataxia y distonía . [25] El modelado numérico de datos experimentales sugiere que, in vivo, el Na +
- K +
la bomba produce largas puntuaciones quiescentes (>> 1 s) a la activación de las neuronas de Purkinje; estos pueden tener un papel computacional. [26] El alcohol inhibe Na +
- K +
Bombea en el cerebelo y es probable que corrompa la computación del cerebelo y la coordinación corporal. [27] [28]
Importancia clínica [ editar ]
En los seres humanos, las células de Purkinje pueden ser dañadas por una variedad de causas: exposición tóxica, por ejemplo al alcohol o litio; enfermedades autoinmunes ; mutaciones genéticas que causan ataxias espinocerebelosas, ataxia por gluten , enfermedad de Unverricht-Lundborg o autismo ; y enfermedades neurodegenerativas que no se sabe que tengan una base genética, como el tipo cerebeloso de atrofia de sistemas múltiples o ataxias esporádicas. [29] [30]
La ataxia del gluten es una enfermedad autoinmune provocada por la ingestión de gluten . [31] La muerte de las células de Purkinje como resultado de la exposición al gluten es irreversible. El diagnóstico precoz y el tratamiento con una dieta sin gluten pueden mejorar la ataxia y prevenir su progresión. [29] [32] Menos del 10% de las personas con ataxia por gluten presentan algún síntoma gastrointestinal, sin embargo, alrededor del 40% tiene daño intestinal. [32] Representa el 40% de las ataxias de origen desconocido y el 15% de todas las ataxias. [32]
La enfermedad neurodegenerativa ataxia espinocerebelosa tipo 1 (SCA1) es causada por una expansión de poliglutamina inestable dentro de la proteína Ataxina 1 . Este defecto en la proteína Ataxin 1 causa deterioro de las mitocondrias en las células de Purkinje, lo que lleva a una degeneración prematura de las células de Purkinje. [33] Como consecuencia, la coordinación motora disminuye y eventualmente se produce la muerte.
Algunos animales domésticos pueden desarrollar una condición en la que las células de Purkinje comienzan a atrofiarse poco después del nacimiento, llamada abiotrofia cerebelar . Puede provocar síntomas como ataxia , temblores de intención, hiperreactividad, falta de reflejo de amenaza , marcha rígida o paso a paso alto, aparente falta de conciencia de la posición del pie (a veces de pie o caminando con el pie doblado) y una incapacidad general para Determinar el espacio y la distancia. [34] Una condición similar conocida como hipoplasia cerebelosaocurre cuando las células de Purkinje no se desarrollan en el útero o mueren antes del nacimiento.
Las condiciones genéticas de la ataxia telangiectasia y la enfermedad de Niemann Pick tipo C, así como el temblor esencial del cerebelo , implican la pérdida progresiva de células de Purkinje. En la enfermedad de Alzheimer, a veces se observa patología espinal, así como pérdida de ramas dendríticas de las células de Purkinje. [35] Las células de Purkinje también pueden ser dañadas por el virus de la rabia a medida que migra desde el sitio de la infección en la periferia al sistema nervioso central.
No hay comentarios:
Publicar un comentario