Las células amacrinas son interneuronas en la retina . [1] Se denominan de las raíces griegas a– ("non"), makr– ("long") y in–("fibre"), debido a sus cortos procesos neuríticos. Las células amacrinas son neuronas inhibitorias, y proyectan sus cenizas dendríticas sobre la capa plexiforme interna (LPI), interactúan con las células ganglionares de la retina y / o las células bipolares .

Las células de la retina de Xenopus se tiñeron para cdk2 / cyclin2 con una flecha roja que indica una célula amacrina. IPL se muestra en blanco.
Estructura [ editar ]
Las células amacrinas operan en la capa nuclear interna (también llamada estrato nuclear interno ) (INL), la segunda capa retiniana sináptica donde las células bipolares y las células ganglionares de la retina forman sinapsis . Hay al menos 33 subtipos diferentes de células amacrinas basadas solo en su morfología y estratificación de dendritas. Al igual que las células horizontales , las célulasamacrinas funcionan lateralmente, pero mientras que las células horizontales están conectadas a la salida de las células de bastón y cono, las células amacrinas afectan la salida de las células bipolares y, a menudo, son más especializadas. Cada tipo de célula amacrina libera uno o varios neurotransmisores donde se conecta con otras células. [2]
A menudo se clasifican por el ancho de su campo de conexión, en qué capa (s) del estrato en la IPL en la que se encuentran y por tipo de neurotransmisor. La mayoría son inhibidores del uso de ácido gamma-aminobutírico o glicina como neurotransmisores .
Tipos [ editar ]
Como se mencionó anteriormente, hay varias formas diferentes de dividir los diferentes tipos de células amacrinas en subtipos.
GABAérgico, glicinérgico o ninguno de los dos : las células amacrinas pueden ser GABAérgicas , glicinérgicas o ninguna dependiendo de qué neurotransmisor inhibidor expresan (GABA, glicina o ninguno). Las células amacrinas GABAérgicas suelen ser células amacrinas de campo amplio y se encuentran en la capa de células ganglionares (GCL) y la capa nuclear interna (INL). Un tipo de célula amacrina GABAérgica que está bastante bien estudiada es la célula amacrina de estallido estelar . Estas células amacrinas se caracterizan generalmente por su expresión de colina acetiltransferasa o ChAT y se sabe que desempeñan un papel en la selectividad de la dirección y la detección del movimiento direccional. [2]La acetilcolina también se libera de estas células amacrinas, pero su función no se conoce completamente. [3] Otro subtipo de células amacrinas GABAérgicas son aquellas que son dopaminérgicas. Estos son todos los que expresan TH y estas células amacrinas modulan la adaptación de la luz y el ritmo circadiano. [2] Estas son células amacrinas ampliamente diseminadas, y liberan difusamente dopamina, mientras que aún liberan GABA y llevan a cabo toda la liberación sináptica normal. [3] Se han observado muchas otras divisiones de células amacrinas GABAérgicas, pero las enumeradas anteriormente son algunas de las más ampliamente investigadas y discutidas.
Las células amacrinas glicinérgicas no están tan caracterizadas como las células amacrinas GABAérgicas. Sin embargo, todas las células amacrinas glicinérgicas están marcadas por el transportador de glicina GlyT1 . Una célula amacrina glicinérgica muy bien caracterizada es la célula amacrina AII . Estas células están presentes en el INL. [2] Una función importante de las células amacrinas AII es que capturan la entrada celular de las células bipolares de varilla y la redistribuyen a células bipolares de cono utilizando las terminaciones sinápticas de las células bipolares de cono como adaptadores [4]
Alrededor del 15% de las células amacrinas no son GABAérgicas ni glicinérgicas. [2] Estas células amacrinas a veces se conocen como células amacrinas nGnG, y se cree que los factores de transcripción que actúan sobre los progenitores deciden el destino de las células amacrinas. Un factor de transcripción que se encontró que se expresaba selectivamente en células amacrinas nGnG es Neurod6 [5]
Longitud de cenizas dendríticas : en función de la extensión y extensión de las cenizas dendríticas, las células amacrinas se pueden clasificar como células amacrinas de campo estrecho (alrededor de 70 micrómetros de diámetro), células amacrinas de campo medio (alrededor de 170 micrómetros de diámetro) y células amacrinas de campo amplio (alrededor de 350 micrómetros de diámetro). [2]Estas diferentes longitudes se prestan a diferentes funciones específicas que las células amacrinas pueden realizar. Las células amacrinas de campo estrecho permiten la comunicación vertical entre diferentes niveles de la retina. También ayudan a crear subunidades funcionales en el campo receptivo de las células ganglionares. Estas células amacrinas de campo estrecho y su superposición en estas subunidades pueden permitir que ciertas células ganglionares detecten pequeñas cantidades de movimiento de un punto muy pequeño en un campo de visión. Un tipo de células de campo estrecho que hace esto es la célula amacrina de estallido estelar. [3]
Las células amacrinas de campo medio también contribuyen a la comunicación vertical en las células de la retina, pero gran parte de su función general es aún desconocida. Debido al hecho de que el tamaño de su árbol dendrítico es bastante similar al de las células ganglionares, podrían confundir el borde del campo visual de las células ganglionares. De manera similar, las células amacrinas de campo amplio son difíciles de investigar e incluso descubrir porque abarcan toda la retina, por lo que no hay muchas de ellas. Sin embargo, debido a su tamaño, una de sus funciones principales es la comunicación lateral dentro de una capa, aunque algunas también se comunican verticalmente entre capas. [3]
Organización [ editar ]
Las células amacrinas y otras células de la interneurona de la retina tienen menos probabilidades de estar cerca de vecinos del mismo subtipo de lo que ocurriría por casualidad, lo que da como resultado "zonas de exclusión" que las separan. Los arreglos de mosaico proporcionan un mecanismo para distribuir cada tipo de célula uniformemente a través de la retina, asegurando que todas las partes del campo visual tengan acceso a un conjunto completo de elementos de procesamiento. [6] Las proteínas transmembrana MEGF10 y MEGF11 tienen funciones críticas en la formación de los mosaicos por las células de amacrina y las células horizontales en ratones. [7]
Función [ editar ]
En muchos casos, el subtipo de la célula amacrina habla de su función (la forma conduce a la función), pero se pueden describir algunas funciones específicas de las células amacrinas de la retina.
- Interceptar células ganglionares de la retina y / o células bipolares en la IPL [2]
- Cree subunidades funcionales dentro de los campos receptivos de muchas células ganglionares
- Contribuir a la comunicación vertical dentro de las capas retinianas.
- Llevar a cabo funciones paracrinas como la liberación de dopamina y acetilcolina [3]
- A través de sus conexiones con otras células de la retina en las sinapsis y la liberación de neurotransmisores, contribuyen a la detección del movimiento direccional, modulan la adaptación de la luz y el ritmo circadiano, [2] y controlan la alta sensibilidad en la visión escotópica a través de conexiones con células bipolares de bastón y cono [4]
Todavía hay mucho por descubrir acerca de todas las diferentes funciones de todas las diferentes células amacrinas. Se cree que las células amacrinas con extensos árboles dendríticos contribuyen al entorno inhibitorio mediante la retroalimentación tanto a nivel de células bipolares como de células ganglionares. En este papel se consideran para complementar la acción de las celdas horizontales.
Es probable que otras formas de células amacrinas desempeñen funciones moduladoras, lo que permite el ajuste de la sensibilidad para la visión fotópica y escotópica . La célula amacrina AII es un mediador de señales de células de varilla en condiciones escotópicas.
Las células espesas son dos tipos de neuronas de segundo orden que se encuentran en la parte anterior del núcleo coclear ventral , el AVCN. Pueden ser globulares o esféricas, dando salidas a diferentes partes del complejo olivar superior .
Estructura [ editar ]
Las células espesas se nombran después de su aparición a los arbustos , que tienen dendritas cortas. Están casi completamente engullidos por las conexiones sinápticas de los bulbos finales de Held . [1]
Función [ editar ]
Debido al gran contacto sináptico de las fibras nerviosas auditivas , el patrón de salida de la célula espesa es casi el mismo que la entrada del nervio auditivo. Las proyecciones de las células arbustivas globulares se extienden a la oliva superior en ambos lados del tronco cerebral, donde dan entrada a las neuronas bipolares. La aceituna superior es un área que se considera importante en el procesamiento de señales binaurales. [1]
Las proyecciones de las células espesas esféricas dan entrada excitadora a las partes laterales y mediales de la oliva superior. Una vez más, su acoplamiento sináptico muy cercano sugiere una parte en el papel del procesamiento de las diferencias de tiempo interaurales .
Las células granulares del cerebelo forman la gruesa capa granular de la corteza cerebelosa y se encuentran entre las neuronas más pequeñas del cerebro. (El término célula granular se usa para varios tipos no relacionados de neuronas pequeñas en varias partes del cerebro). Las células granulares cerebelosas también son las neuronas más numerosas en el cerebro: en los humanos, las estimaciones de su número total promedian alrededor de 50 mil millones, lo que significa que constituyen alrededor de 3/4 de las neuronas del cerebro.
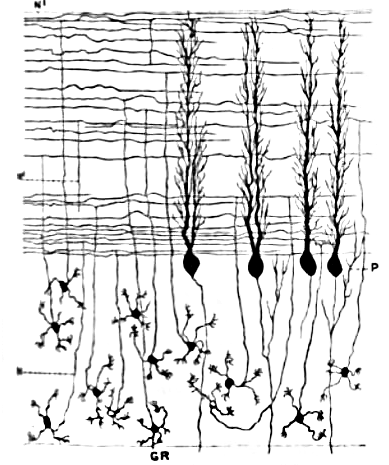
Estructura [ editar ]
Los cuerpos celulares se empaquetan en una gruesa capa granular en la parte inferior de la corteza cerebelosa. Una célula de gránulo emite solo de cuatro a cinco dendritas, cada una de las cuales termina en un agrandamiento llamado garra dendrítica . [1] Estas ampliaciones son sitios de entrada excitadora de fibras musgosas y entrada inhibitoria de las células de Golgi.
Los delgados y no mielinizados axones de las células granulares se elevan verticalmente a la capa superior (molecular) de la corteza, donde se dividen en dos, con cada rama viajando horizontalmente para formar una fibra paralela ; La división de la rama vertical en dos ramas horizontales da lugar a una forma de "T" distintiva. Una fibra paralela recorre un promedio de 3 mm en cada dirección desde la división, para una longitud total de aproximadamente 6 mm (aproximadamente 1/10 del ancho total de la capa cortical). [1] A medida que avanzan, las fibras paralelas pasan a través de los árboles dendríticos de las células de Purkinje, en contacto con uno de cada 3-5 que pasan, formando un total de 80-100 conexiones sinápticas con espinas dendríticas de células de Purkinje. [1] Las células granulares utilizan glutamato como su neurotransmisor, y por lo tanto ejercen efectos excitadores sobre sus objetivos.
Desarrollo [ editar ]
En el desarrollo normal, la señalización endógena de Sonic hedgehog estimula la rápida proliferación de progenitores de neuronas de gránulos cerebelosos (CGNP) en la capa externa de gránulos (EGL). El desarrollo del cerebelo se produce durante la embriogénesis tardía y el período postnatal temprano, con la proliferación de CGNP en el pico de EGL durante el desarrollo temprano (P7, día 7 postnatal, en el ratón). [2] Como las CGNP se diferencian finalmente en células granulares del cerebelo (también llamadas neuronas granulares del cerebelo, CGN), migran a la capa interna del gránulo (IGL), formando el cerebelo maduro (por P20, día 20 postnatal en el ratón). [2] Las mutaciones que activan de manera anormal la señalización de Sonic hedgehog predisponen al cáncer del cerebelo (meduloblastoma ) en humanos con síndrome de Gorlin y en modelos de ratones diseñados genéticamente . [3] [4]
Función [ editar ]
Las células de los gránulos reciben todo su aporte de las fibras musgosas, pero las superan en número de 200 a 1 (en humanos). Por lo tanto, la información en el estado de actividad de la población de células granulosas es la misma que la información en las fibras musgosas, pero recodificada de una manera mucho más amplia. Debido a que las células granulares son tan pequeñas y tan densamente compactadas, ha sido muy difícil registrar su actividad de espiga en los animales que se comportan, por lo que hay poca información para usar como base de la teorización. El concepto más popular de su función fue propuesto por David Marr , quien sugirió que podrían codificar combinaciones.De insumos de fibra de musgo. La idea es que con cada celda de gránulo que recibe entradas de solo 4–5 fibras musgosas, una celda de gránulo no respondería si solo una de sus entradas estuviera activa, pero respondería si más de una estuviera activa. Este esquema de "codificación combinatoria" potencialmente permitiría al cerebelo hacer distinciones mucho más finas entre los patrones de entrada que lo que permitirían las fibras musgosas solas.
No hay comentarios:
Publicar un comentario